

At5PTase13 modulates cotyledon vein development through regulating auxin homeostasis
- PMID: 16299182
- PMCID: PMC1310551
- DOI: 10.1104/pp.105.067140
At5PTase13 modulates cotyledon vein development through regulating auxin homeostasis
Abstract
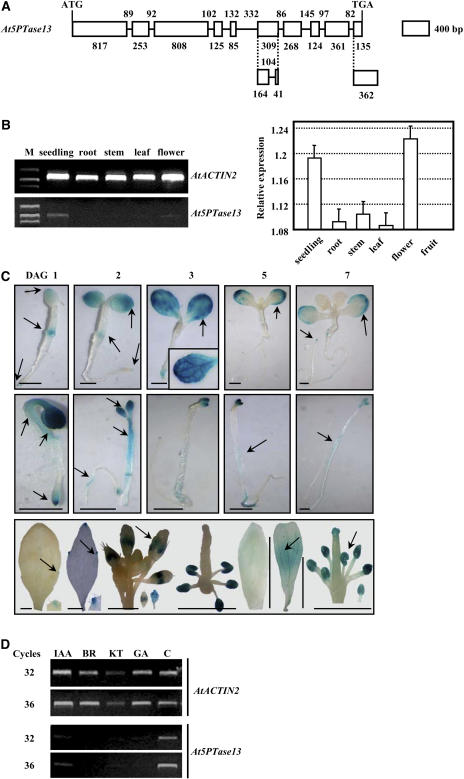
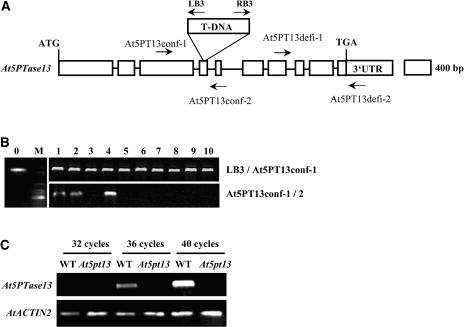
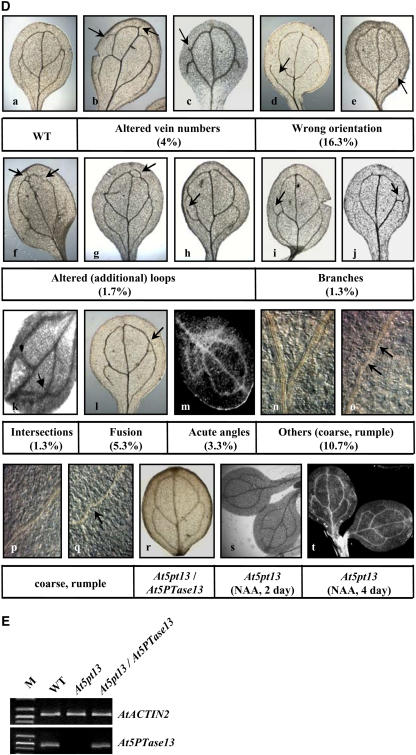
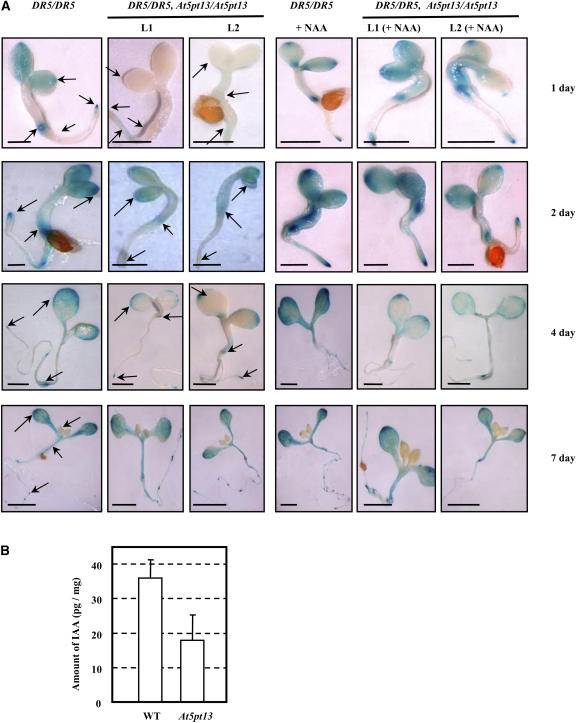
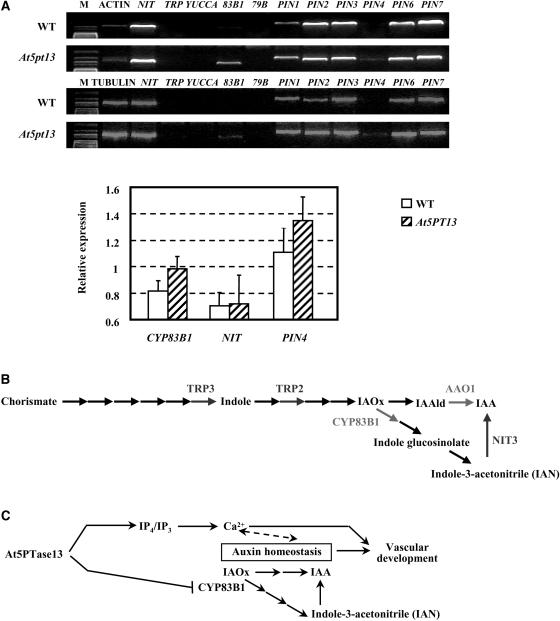
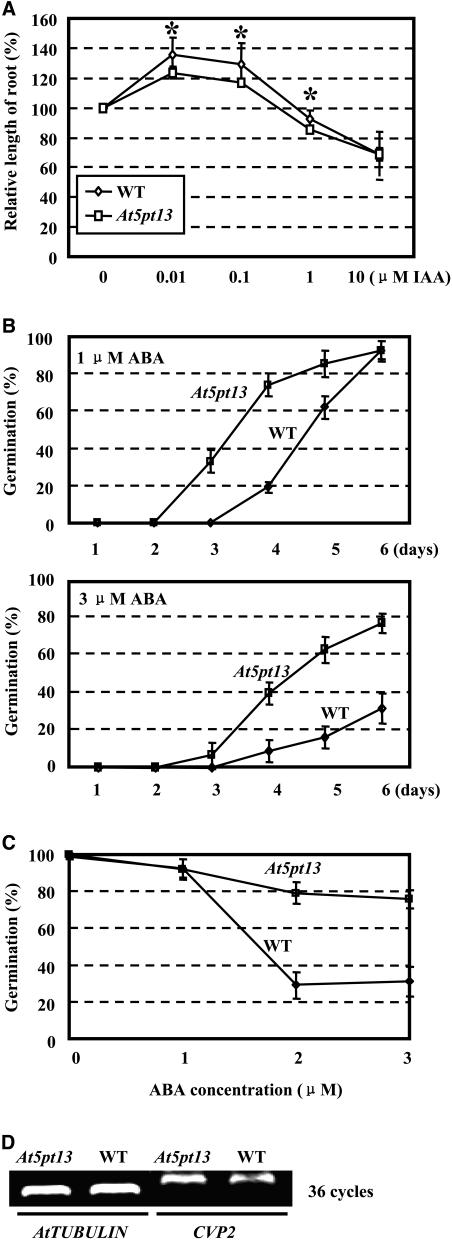
Phosphatidylinositol signaling pathway and the relevant metabolites are known to be critical to the modulation of different aspects of plant growth, development, and stress responses. Inositol polyphosphate 5-phosphatase is a key enzyme involved in phosphatidylinositol metabolism and is encoded by an At5PTase gene family in Arabidopsis thaliana. A previous study shows that At5PTase11 mediates cotyledon vascular development probably through the regulation of intracellular calcium levels. In this study, we provide evidence that At5PTase13 modulates the development of cotyledon veins through its regulation of auxin homeostasis. A T-DNA insertional knockout mutant, At5pt13-1, showed a defect in development of the cotyledon vein, which was rescued completely by exogenous auxin and in part by brassinolide, a steroid hormone. Furthermore, the mutant had reduced auxin content and altered auxin accumulation in seedlings revealed by the DR5:beta-glucuronidase fusion construct in seedlings. In addition, microarray analysis shows that the transcription of key genes responsible for auxin biosynthesis and transport was altered in At5pt13-1. The At5pt13-1 mutant was also less sensitive to auxin inhibition of root elongation. These results suggest that At5PTase13 regulates the homeostasis of auxin, a key hormone controlling vascular development in plants.
Figures
References
-
- Alfandari D, Darribere T (1994) A simple PCR method for screening cDNA libraries. PCR Methods Appl 4: 46–49 - PubMed
-
- Bartel B, LeClere S, Magidin M, Zolman BK (2001) Inputs to the active indole-3-acetic acid pool: de novo synthesis, conjugate hydrolysis, and indole-3-butyric acid β-oxidation. J Plant Growth Regul 20: 198–216
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
