

Crosstalk in G protein-coupled receptors: changes at the transmembrane homodimer interface determine activation
- PMID: 16301531
- PMCID: PMC1287488
- DOI: 10.1073/pnas.0508950102
Crosstalk in G protein-coupled receptors: changes at the transmembrane homodimer interface determine activation
Abstract
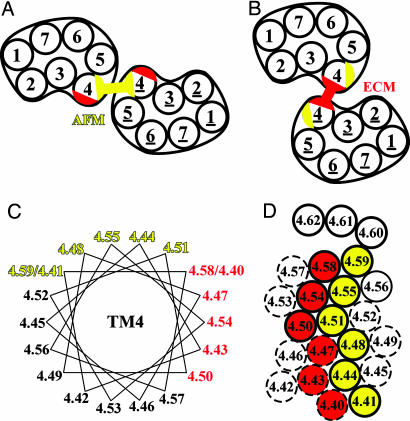
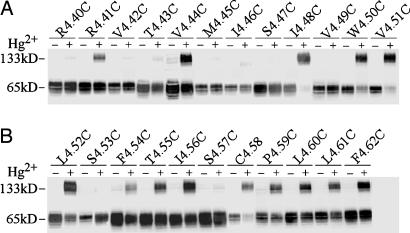
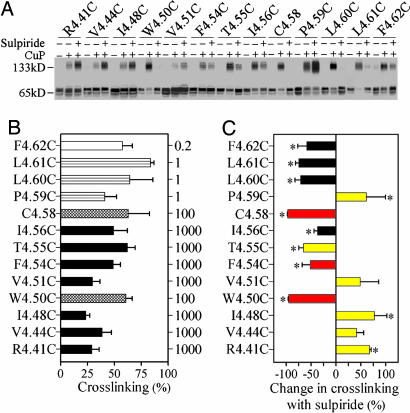
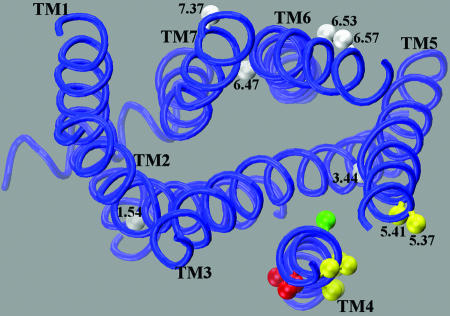
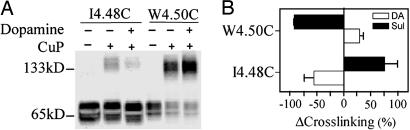
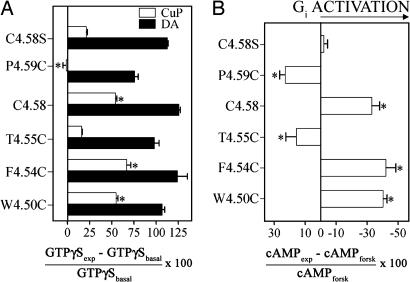
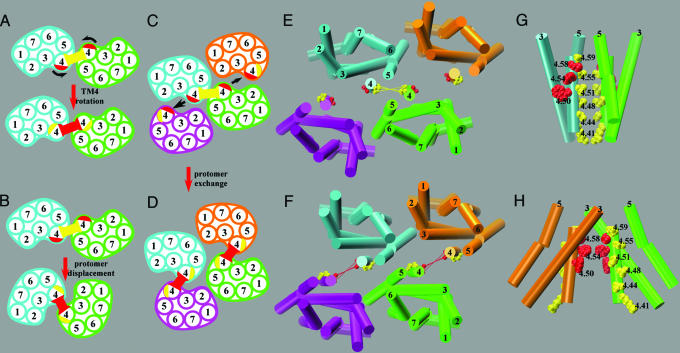
Functional crosstalk between G protein-coupled receptors in a homo- or heterodimeric assembly likely involves conformational changes at the dimer interface, but the nature of this interface is not yet established, and the dynamic changes have not yet been identified. We have mapped the homodimer interface in the dopamine D2 receptor over the entire length of the fourth transmembrane segment (TM4) by crosslinking of substituted cysteines. Their susceptibilities to crosslinking are differentially altered by the presence of agonists and inverse agonists. The TM4 dimer interface in the inverse agonist-bound conformation is consistent with the dimer of the inactive form of rhodopsin modeled with constraints from atomic force microscopy. Crosslinking of a different set of cysteines in TM4 was slowed by inverse agonists and accelerated in the presence of agonists; crosslinking of the latter set locks the receptor in an active state. Thus, a conformational change at the TM4 dimer interface is part of the receptor activation mechanism.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
