

A point mutation within the replicase gene differentially affects coronavirus genome versus minigenome replication
- PMID: 16306572
- PMCID: PMC1316003
- DOI: 10.1128/JVI.79.24.15016-15026.2005
A point mutation within the replicase gene differentially affects coronavirus genome versus minigenome replication
Abstract
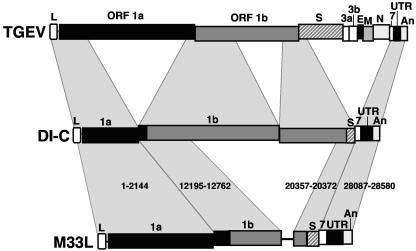
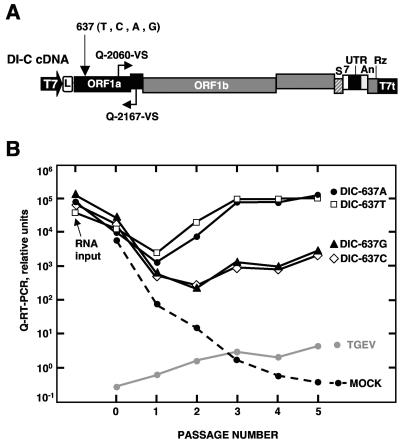
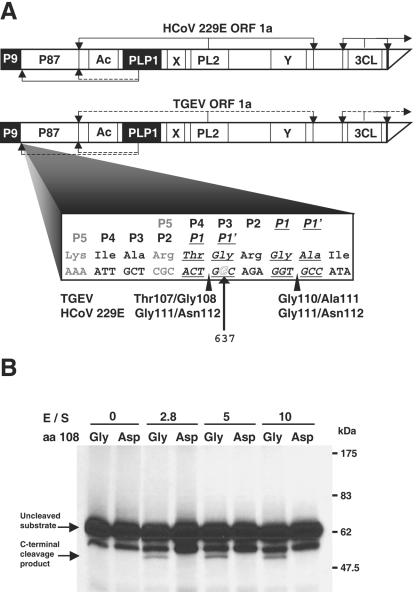
During the construction of the transmissible gastroenteritis virus (TGEV) full-length cDNA clone, a point mutation at position 637 that was present in the defective minigenome DI-C was maintained as a genetic marker. Sequence analysis of the recovered viruses showed a reversion at this position to the original virus sequence. The effect of point mutations at nucleotide 637 was analyzed by reverse genetics using a TGEV full-length cDNA clone and cDNAs from TGEV-derived minigenomes. The replacement of nucleotide 637 of TGEV genome by a T, as in the DI-C sequence, or an A severely affected virus recovery from the cDNA, yielding mutant viruses with low titers and small plaques compared to those of the wild type. In contrast, T or A at position 637 was required for minigenome rescue in trans by the helper virus. No relationship between these observations and RNA secondary-structure predictions was found, indicating that mutations at nucleotide 637 most likely had an effect at the protein level. Nucleotide 637 occupies the second codon position at amino acid 108 of the pp1a polyprotein. This position is predicted to map in the N-terminal polyprotein papain-like proteinase (PLP-1) cleavage site at the p9/p87 junction. Replacement of G-637 by A, which causes a drastic amino acid change (Gly to Asp) at position 108, affected PLP-1-mediated cleavage in vitro. A correlation was found between predicted cleaving and noncleaving mutations and efficient virus rescue from cDNA and minigenome amplification, respectively.
Figures
Similar articles
-
The spike protein of transmissible gastroenteritis coronavirus controls the tropism of pseudorecombinant virions engineered using synthetic minigenomes.Adv Exp Med Biol. 1998;440:207-14. doi: 10.1007/978-1-4615-5331-1_25. Adv Exp Med Biol. 1998. PMID: 9782282
-
Replication and packaging of transmissible gastroenteritis coronavirus-derived synthetic minigenomes.J Virol. 1999 Feb;73(2):1535-45. doi: 10.1128/JVI.73.2.1535-1545.1999. J Virol. 1999. PMID: 9882359 Free PMC article.
-
Mutations in the GDD motif of rubella virus putative RNA-dependent RNA polymerase affect virus replication.Virology. 2001 Jul 5;285(2):322-31. doi: 10.1006/viro.2001.0939. Virology. 2001. PMID: 11437666
-
Development of mouse hepatitis virus and SARS-CoV infectious cDNA constructs.Curr Top Microbiol Immunol. 2005;287:229-52. doi: 10.1007/3-540-26765-4_8. Curr Top Microbiol Immunol. 2005. PMID: 15609514 Free PMC article. Review.
-
Expression of transcriptional units using transmissible gastroenteritis coronavirus derived minigenomes and full-length cDNA clones.Adv Exp Med Biol. 2001;494:447-51. doi: 10.1007/978-1-4615-1325-4_65. Adv Exp Med Biol. 2001. PMID: 11774506 Review. No abstract available.
Cited by
-
Structure of nonstructural protein 1 from SARS-CoV-2.bioRxiv [Preprint]. 2020 Nov 3:2020.11.03.366757. doi: 10.1101/2020.11.03.366757. bioRxiv. 2020. Update in: J Virol. 2021 Jan 28;95(4):e02019-20. doi: 10.1128/JVI.02019-20. PMID: 33173873 Free PMC article. Updated. Preprint.
-
Biochemical aspects of coronavirus replication.Adv Exp Med Biol. 2006;581:13-24. doi: 10.1007/978-0-387-33012-9_2. Adv Exp Med Biol. 2006. PMID: 17037498 Free PMC article. Review. No abstract available.
-
Molecular characterization of a Chinese vaccine strain of transmissible gastroenteritis virus: mutations that may contribute to attenuation.Virus Genes. 2010 Jun;40(3):403-9. doi: 10.1007/s11262-010-0467-6. Epub 2010 Mar 13. Virus Genes. 2010. PMID: 20229183 Free PMC article.
-
Novel beta-barrel fold in the nuclear magnetic resonance structure of the replicase nonstructural protein 1 from the severe acute respiratory syndrome coronavirus.J Virol. 2007 Apr;81(7):3151-61. doi: 10.1128/JVI.01939-06. Epub 2007 Jan 3. J Virol. 2007. PMID: 17202208 Free PMC article.
-
Transmissible Gastroenteritis Virus (TGEV) and Porcine Respiratory Coronavirus (PRCV): Epidemiology and Molecular Characteristics-An Updated Overview.Viruses. 2025 Mar 28;17(4):493. doi: 10.3390/v17040493. Viruses. 2025. PMID: 40284936 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
