

Episomal maintenance of plasmids with hybrid origins in mouse cells
- PMID: 16306599
- PMCID: PMC1316011
- DOI: 10.1128/JVI.79.24.15277-15288.2005
Episomal maintenance of plasmids with hybrid origins in mouse cells
Abstract
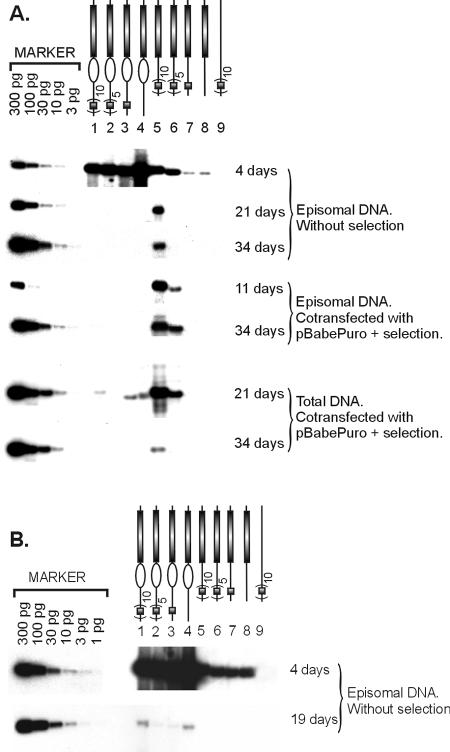
Bovine papillomavirus type 1 (BPV1), Epstein-Barr virus (EBV), and human herpesvirus 8 genomes are stably maintained as episomes in dividing host cells during latent infection. The mitotic segregation/partitioning function of these episomes is dependent on single viral protein with specific DNA-binding activity and its multimeric binding sites in the viral genome. In this study we show that, in the presence of all essential viral trans factors, the segregation/partitioning elements from both BPV1 and EBV can provide the stable maintenance function to the mouse polyomavirus (PyV) core origin plasmids but fail to do so in the case of complete PyV origin. Our study is the first which follows BPV1 E2- and minichromosome maintenance element (MME)-dependent stable maintenance function with heterologous replication origins. In mouse fibroblast cell lines expressing PyV large T antigen (LT) and either BPV1 E2 or EBV EBNA1, the long-term episomal replication of plasmids carrying the PyV minimal origin together with the MME or family of repeats (FR) element can be monitored easily for 1 month under nonselective conditions. Our data demonstrate clearly that the PyV LT-dependent replication function and the segregation/partitioning function of the BPV1 or EBV are compatible in certain, but not all, configurations. The quantitative analysis indicates a loss rate of 6% per cell, doubling in the case of MME-dependent plasmids, and 13% in the case of FR-dependent plasmids in nonselective conditions. Our data clearly indicate that maintenance functions from different viruses are principally interexchangeable and can provide a segregation/partitioning function to different heterologous origins in a variety of cells.
Figures






References
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, and K. Struhl (ed.). 1998. Current protocols in molecular biology. John Wiley & Sons, Inc., New York, N.Y.
-
- Ballestas, M. E., P. A. Chatis, and K. M. Kaye. 1999. Efficient persistence of extrachromosomal KSHV DNA mediated by latency-associated nuclear antigen. Science 284:641-644. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
