

doi: 10.1128/JVI.79.24.15567-15572.2005.
Restriction of human immunodeficiency virus type 1 by TRIM-CypA occurs with rapid kinetics and independently of cytoplasmic bodies, ubiquitin, and proteasome activity
Affiliations
- PMID: 16306627
- PMCID: PMC1316013
- DOI: 10.1128/JVI.79.24.15567-15572.2005
Item in Clipboard
Restriction of human immunodeficiency virus type 1 by TRIM-CypA occurs with rapid kinetics and independently of cytoplasmic bodies, ubiquitin, and proteasome activity
J Virol.
2005 Dec.
Abstract
TRIM-CypA is an owl monkey-specific variant of the retrovirus restriction factor TRIM5alpha. Here, we exploit its modular domain organization and cyclosporine sensitivity to probe the kinetics and mechanism of TRIM5-mediated restriction. Time of addition/withdrawal experiments reveal that inhibition of incoming human immunodeficiency virus type 1 capsids by TRIM-CypA occurs within minutes of their delivery to the target cell cytoplasm. However, while TRIM-CypA restriction is partly dependent on a RING domain, restriction occurs independently of the ubiquitin/proteasome system. Moreover, tagged TRIM-CypA proteins can be fully active as restriction factors without forming cytoplasmic bodies.
Figures

Kinetic analysis of HIV-1 restriction by TRIM-CypA. (A) Experimental strategy. HXB-enveloped HIV-1 particles were bound to OMK-CD4/CXCR4 cells, and infection was initiated by a temperature shift. Infection events were quantitated 48 h after the temperature shift by enumerating GFP-positive cells using fluorescence-activated cell sorter analysis. The time taken to acquire resistance to subsequently applied inhibitors of entry and reverse transcription and the opposing effects of CsA application and removal were measured. (B) Quantitation of infection events (percent GFP-positive cells at 48 h after the temperature shift) following timed addition of an entry inhibitor, dextran sulfate (Dex S) or UC781, a nonnucleoside reverse transcriptase (RT) inhibitor (dashed lines). Addition of either inhibitor at the time of the temperature shift (0 h) reduced infection to undetectable levels (>1,000-fold). Alternatively, TRIM-CypA restriction was activated or inhibited by the withdrawal or addition of CsA (solid lines). (C) A portion of the data in panel B is shown, except that only early time points are plotted on an expanded time axis and infection events are plotted on a linear scale as a percentage of their maximum uninhibited level.

Cytoplasmic body formation by TRIM-CypA occurs only at high expression levels and is not required for restriction. (A) OMK cells, unmodified MDTF cells (none), or cloned MDTF cells stably expressing TRIM-CypA-HA or TRIM-CypA-CFP (clones 1 and 2) using LNCX2-derived retroviral vectors were infected with VSV-G-pseudotyped HIV-1 or HIV(SIV CA) vectors, as indicated. The percentage of infected (GFP-positive) cells as determined by FACS analysis is plotted, and results of one representative of two independent experiments are shown. (B) Comparison of the expression levels of TRIM-CypA-HA in untreated and sodium butyrate-treated MDTF cells. Cell lysates were fourfold serially diluted (beginning at 105 cell equivalents per lane) and subjected to Western blot (WB) analysis using an anti-HA (α-HA) tag monoclonal antibody. (C) Same as panel B except that TRIM-CypA-CFP (clones 1 and 2) we analyzed using an anti-GFP (αGFP) antibody. Note that the anti-HA antibody permits more-sensitive detection than the anti-GFP antibody; therefore, the signals in panels B and C cannot be meaningfully compared with each other. (D) Localization of TRIM-CypA-HA and TRIMCyp-CFP in untreated cells (left) and cells treated with 10 mM sodium butyrate for 16 h prior to fixation (right). TRIM-Cyp-HA localization in fixed and permeabilized cells was determined using anti-HA primary antibodies and Alexa-594-congugated secondary antibodies. TRIM-CypA-CFP localization was determined in fixed but otherwise unmanipulated cells. A single 0.25-μm deconvolved optical section is shown in each image, and selection of three representative images is shown for each cell line/condition. IF, immunofluorescence.
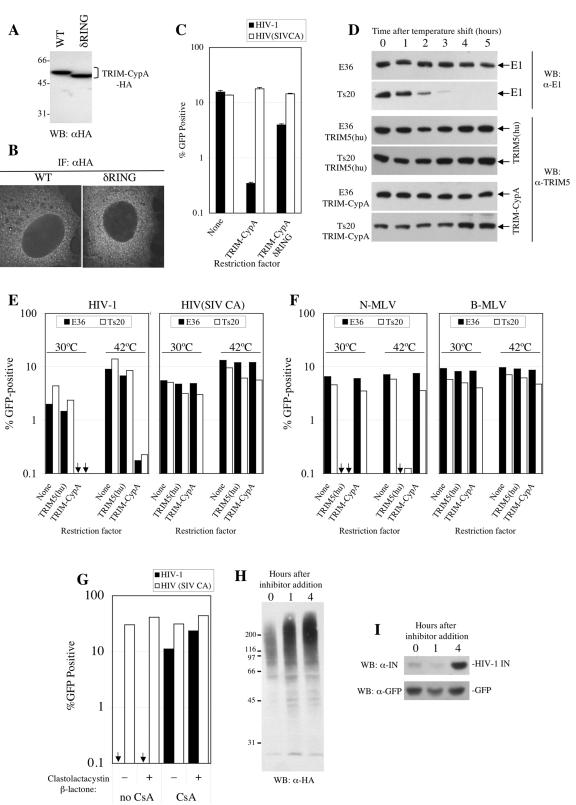
Requirement for the RING domain but not ubiquitin-activating (E1) enzyme or proteasome activity in TRIM-CypA restriction. (A) Expression of TRIM-CypA and δRING TRIM-CypA (lacking residues 2 to 60). Lysates from G418-resistant pools of Mus dunni (MDTF) cells transduced with LNCX2-derived retroviral vectors expressing the indicated proteins were analyzed by Western blot (WB) analysis using an anti-HA (α-HA) monoclonal antibody. WT, wild type. (B) Localization of HA-tagged TRIM-CypA or δRING TRIM-CypA. The same MDTF cells analyzed in panel A were subjected to immunofluorescence (IF) analysis as described for Fig. 2D. (C) Restriction activity of TRIM-CypA and δRING TRIM-CypA in MDTF cells. Unmodified MDTF cells or MDTF cells stably expressing HA-tagged TRIM-CypA or δRING TRIM-CypA were infected with VSV-G-pseudotyped HIV-1 or HIV(SIV CA) vectors, as indicated. The percentage of infected (GFP-positive) cells as determined by FACS analysis 48 h later is plotted. (D) Specific E1 removal from Ts20 cells. Unmodified E36 or Ts20 cells or cells engineered to express either TRIM5α or TRIM-CypA were cultured at 30°C (time zero) and subsequently shifted to 42°C for the indicated number of hours. Cell lysates were subjected to Western blotting with antibodies specific for ubiquitin-activating (E1) enzyme or TRIM5, as indicated. (E) Lack of requirement of E1 in TRIM-CypA restriction. Unmodified Ts20 or control E36 cells (None) or cells stably expressing either TRIM-CypA or human TRIM5α [TRIM5(hu)] were incubated at the indicated temperatures for 3 h prior to and 2 h subsequent to infection with a VSV-G-pseudotyped HIV-1 or HIV(SIV CA) vector, as indicated, before being washed and returned to 30°C. The percentages of infected (GFP-positive) cells as determined by FACS analysis 48 h later are plotted. (F) The same experiment as described for panel E was performed except that the various E36- and Ts20-derived cell lines were infected with TRIM5(hu)-restricted N-MLV or unrestricted B-tropic MLV vectors, as indicated. (G) Lack of effect of proteasome inhibition on TRIM-CypA restriction in OMK cells. Cells were infected with VSV-G-pseudotyped HIV-1 or HIV(SIV CA) vectors as indicated in the presence (+) or absence (−) of 10 μM clastolactacystin-β-lactone for 1 h prior, 2 h during, and 1 h after exposure to the inoculum. Infections were done under HIV-1-restricting conditions (no CsA) or in the absence of restriction (with CsA). (H, I) Effect of clastolactacystin-β-lactone on HA-ubiquitin conjugate accumulation or HIV-1 integrase expression. OMK cells were transfected with an HA ubiquitin expression plasmid (H) or pEGFP*IRES-Ub-F-IN CTE (14), which expresses both GFP and an HIV-I integrase bearing an N-terminal Phe residue (I). Cells were treated with clastolactacystin-β-lactone for the indicated number of hours, and lysates were subjected to Western blot analysis with antibodies to the HA tag (H), HIV-1 integrase (IN), or GFP (I).
References
-
- Bieniasz, P. D. 2004. Intrinsic immunity: a front-line defense against viral attack. Nat. Immunol. 5:1109-1115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
