

Isolation and characterization of a protochordate histocompatibility locus
- PMID: 16306984
- PMCID: PMC1401502
- DOI: 10.1038/nature04150
Isolation and characterization of a protochordate histocompatibility locus
Abstract
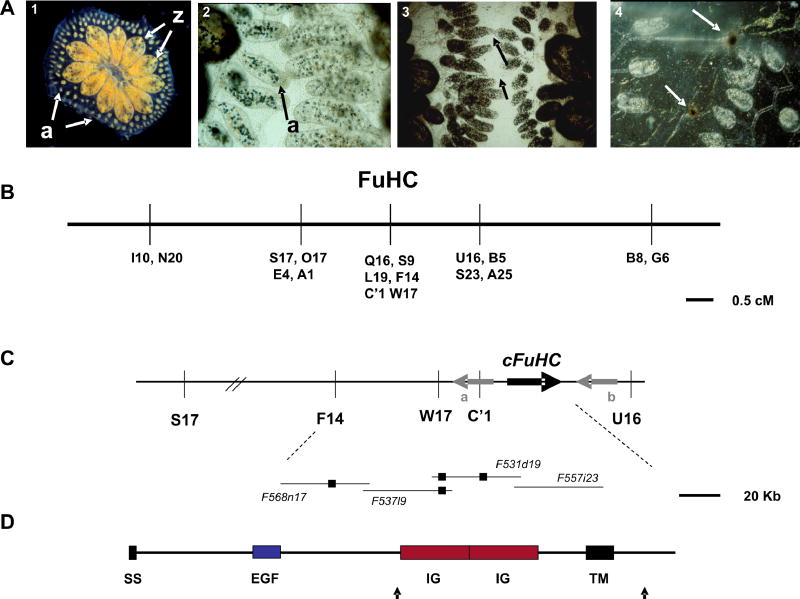
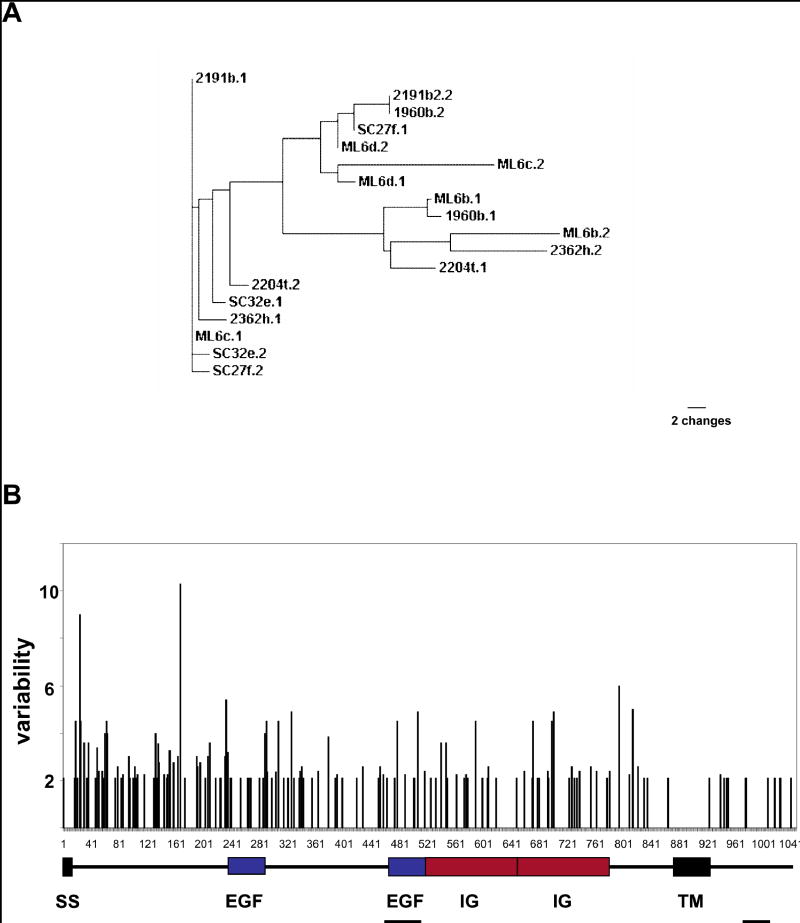
Histocompatibility--the ability of an organism to distinguish its own cells and tissue from those of another--is a universal phenomenon in the Metazoa. In vertebrates, histocompatibility is a function of the immune system controlled by a highly polymorphic major histocompatibility complex (MHC), which encodes proteins that target foreign molecules for immune cell recognition. The association of the MHC and immune function suggests an evolutionary relationship between metazoan histocompatibility and the origins of vertebrate immunity. However, the MHC of vertebrates is the only functionally characterized histocompatibility system; the mechanisms underlying this process in non-vertebrates are unknown. A primitive chordate, the ascidian Botryllus schlosseri, also undergoes a histocompatibility reaction controlled by a highly polymorphic locus. Here we describe the isolation of a candidate gene encoding an immunoglobulin superfamily member that, by itself, predicts the outcome of histocompatibility reactions. This is the first non-vertebrate histocompatibility gene described, and may provide insights into the evolution of vertebrate adaptive immunity.
Figures

Comment in
-
Histocompatibility: colonial match and mismatch.Nature. 2005 Nov 24;438(7067):437-9. doi: 10.1038/438437a. Nature. 2005. PMID: 16306979 No abstract available.
References
-
- Flajnik, M.F., Miller, K. & Dupasquier, L. Evolution of the immune system in: W.E. Paul, Editors, Fundamental Immunology (5th ed), Lippincott, Williams & Wilkins, Philadelphia pp. 519–570. (2003),
-
- Dupasquier L, Zucchetti I, De Santis R. Immunoglobulin superfamily receptors in protochordates : before RAG time. Immunol Rev. 2004;198:233–248. - PubMed
-
- Oka H, Watanabe H. Problems of colony specificity in compound ascidians. Bull Biol Stn Asamushi. 1960;10:153–155.
-
- Sabbadin A. Le basi genetiche dell capacita di fusione fra colonie in Botryllus schlosseri (Ascidiacea) Atti Accad Na Lincei Rend. 1962;32:1031–1035.
-
- Scofield VL, Schlumpberger JM, West LA, Weissman IL. Protochordate allorecognition is controlled by an MHC-like gene system. Nature. 1982;295:499–502. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
