

Review
doi: 10.1016/j.virusres.2005.10.012.
Epub 2005 Nov 22.
End-to-end communication in the modulation of translation by mammalian RNA viruses
Affiliations
- PMID: 16307817
- PMCID: PMC7172311
- DOI: 10.1016/j.virusres.2005.10.012
Item in Clipboard
Review
End-to-end communication in the modulation of translation by mammalian RNA viruses
Virus Res.
2006 Jul.
Abstract
A 5'-3' end interaction leading to stimulation of translation has been described for many cellular and viral mRNAs. Enhancement of viral translational efficiency mediated by 5' and 3' untranslated regions (UTRs) has been shown to occur via RNA-RNA interactions or novel RNA-protein interactions. Mammalian RNA viruses make use of end-to-end communication in conjunction with both viral and cellular factors to regulate multiple processes including translation initiation and the switch between translation and RNA synthesis during the viral lifecycle.
Figures
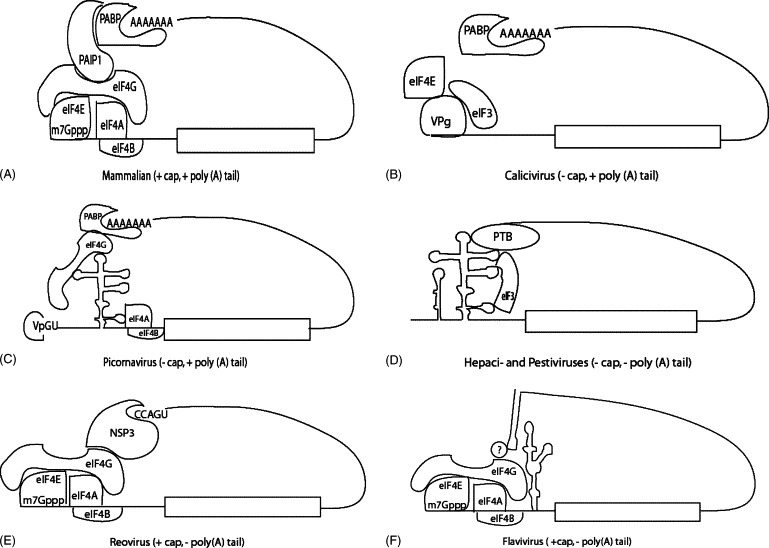
Mechanisms of translation initiation. (A) Mammalian mRNA, (B) calicivirus RNA, (C) picornavirus RNA. Shown is the VPg moiety that is covalently bound to the 5′ end of the genomic RNA, (D) hepaci- and pestivirus RNA, (E) reovirus RNA, and (F) flavivirus RNA.
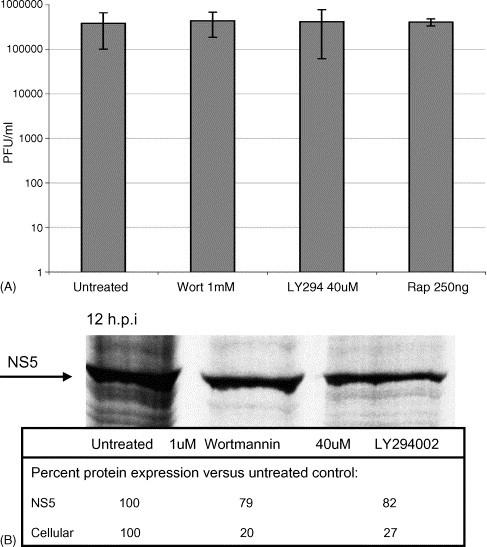
DENV replication and translation are not affected by inhibitors of cap-dependent translation, (A) DENV is replicated in the presence of rapamycin, LY294002 or wortmannin. Cells were exposed to DENV for 1 h in the presence of inhibitors, washed, and incubated with rapamycin (250 ng/ml), LY294002 (40 μM) or wortmannin (1 μM) for 24 h. Supernatants were collected and titered by plaque assay (PFU/ml), (B) DENV RNA is translated in the presence of LY294002 or wortmannin. Cells were prepared as described above. At 12 h post-infection, cells were metabolically labeled and analyzed by SDS-PAGE. ImageQuant software (Molecular Dynamics) was used to quantify the DENV protein NS5 (arrow) and representative cellular proteins.
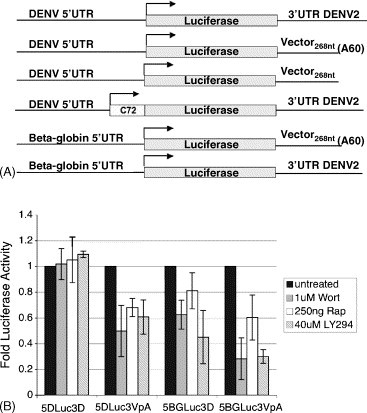
Translation of DENV reporter RNA is not affected by inhibitors of cap-dependent translation (A) DENV RNA reporter constructs. RNA reporter constructs were generated containing either the DENV-2 5′UTR plus or minus the first 72 nt of the DENV coding region or the human β-globin (βg) 5′UTR fused to the firefly luciferase (Luc) gene, followed by either vector sequence, the DENV-2 3′UTR or vector sequence plus a 60-mer poly(A) tail. RNA transcribed in vitro from these constructs using T7 polymerase was transfected into cells, (B) m7G-capped DENV RNA reporter constructs are translated in BHK cells treated with rapamycin, LY294002 or wortmannin. Equal amounts of in vitro transcripts of the constructs described above were capped with m7GpppN and transfected into BHK cells. One hour post-transfection, cells were washed and 250 ng/ml rapamycin, 40 μM LY294002 or 1 μM wortmannin were added. Luc activity was assayed after 12 h by luminometry.
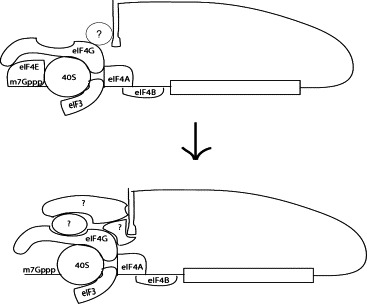
Model of the reorganization of RNP complexes at the 5′ and 3′UTRs of the flavivirus genome under conditions of limiting eIF4E.
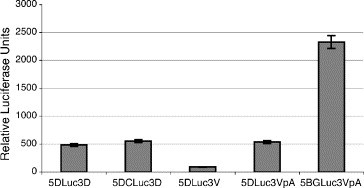
DENV translation is independent of the CS. Equal amounts of m7GpppA-capped in vitro transcripts of the reporter constructs containing the DENV 5′ and 3′UTRs plus or minus the CS were transfected into cells, along with control reporter RNAs (see Fig. 2A). Cells were washed 1 h post-transfection, and Luc activity was assayed after 8 h by luminometry.
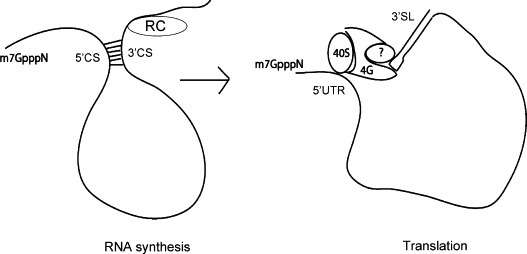
Model depicting the switch between translation and replication of the flavivirus genome mediated through the architectural remodeling of viral RNA.
References
-
- Alvarez D.E., De Lella Ezcurra A.L., Fucito S., Gamarnik A.V. Role of RNA structures present at the 3′UTR of dengue virus on translation, RNA synthesis, and viral replication. Virology. 2005;339:200–212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
