

Molecular structure of a 9-MDa icosahedral pyruvate dehydrogenase subcomplex containing the E2 and E3 enzymes using cryoelectron microscopy
- PMID: 16308322
- PMCID: PMC1647297
- DOI: 10.1074/jbc.M504363200
Molecular structure of a 9-MDa icosahedral pyruvate dehydrogenase subcomplex containing the E2 and E3 enzymes using cryoelectron microscopy
Abstract
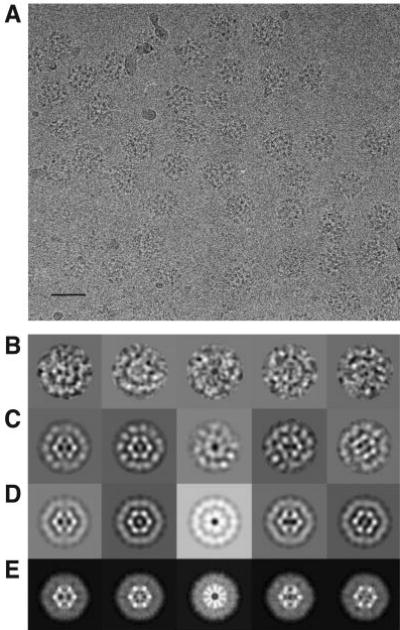
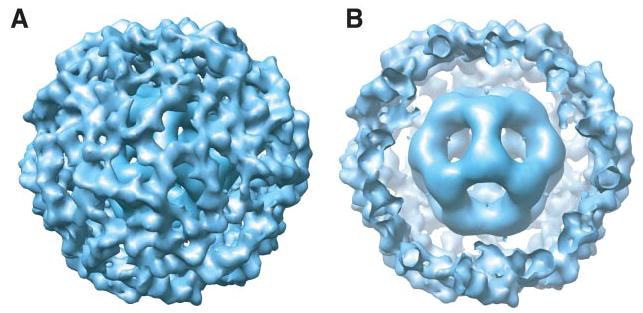
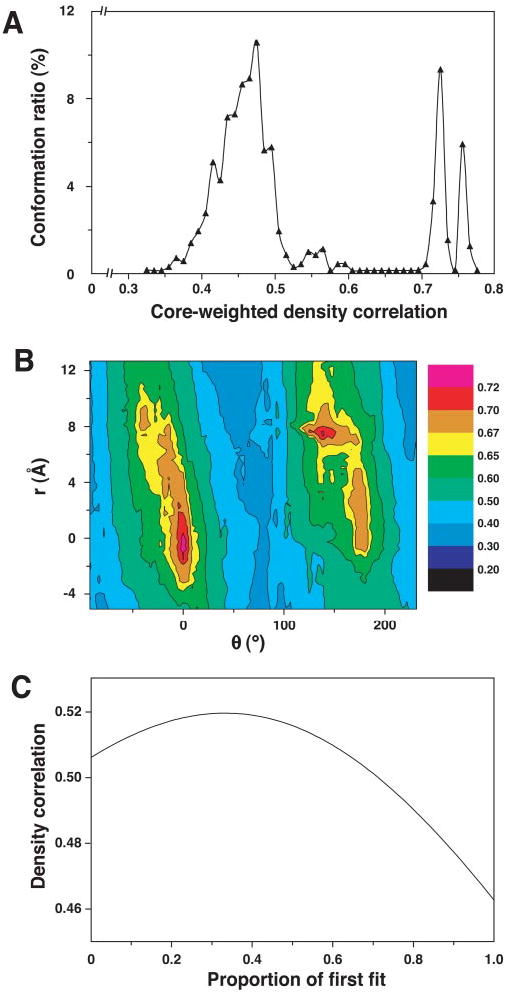
The pyruvate dehydrogenase multienzyme complexes are among the largest multifunctional catalytic machines in cells, catalyzing the production of acetyl CoA from pyruvate. We have previously reported the molecular architecture of an 11-MDa subcomplex comprising the 60-mer icosahedral dihydrolipoyl acetyltransferase (E2) decorated with 60 copies of the heterotetrameric (alpha(2)beta(2)) 153-kDa pyruvate decarboxylase (E1) from Bacillus stearothermophilus (Milne, J. L. S., Shi, D., Rosenthal, P. B., Sunshine, J. S., Domingo, G. J., Wu, X., Brooks, B. R., Perham, R. N., Henderson, R., and Subramaniam, S. (2002) EMBO J. 21, 5587-5598). An annular gap of approximately 90 A separates the acetyltransferase catalytic domains of the E2 from an outer shell formed of E1 tetramers. Using cryoelectron microscopy, we present here a three-dimensional reconstruction of the E2 core decorated with 60 copies of the homodimeric 100-kDa dihydrolipoyl dehydrogenase (E3). The E2E3 complex has a similar annular gap of approximately 75 A between the inner icosahedral assembly of acetyltransferase domains and the outer shell of E3 homodimers. Automated fitting of the E3 coordinates into the map suggests excellent correspondence between the density of the outer shell map and the positions of the two best fitting orientations of E3. As in the case of E1 in the E1E2 complex, the central 2-fold axis of the E3 homodimer is roughly oriented along the periphery of the shell, making the active sites of the enzyme accessible from the annular gap between the E2 core and the outer shell. The similarities in architecture of the E1E2 and E2E3 complexes indicate fundamental similarities in the mechanism of active site coupling involved in the two key stages requiring motion of the swinging lipoyl domain across the annular gap, namely the synthesis of acetyl CoA and regeneration of the dithiolane ring of the lipoyl domain.
Figures




References
-
- Patel MS, Harris RA. FASEB J. 1995;9:1164–1172. - PubMed
-
- Yeaman SJ, Kirby JA, Jones DEJ. Immunol Rev. 2000;174:238–249. - PubMed
-
- Reed LJ, Hackert ML. J Biol Chem. 1990;265:8971–8974. - PubMed
-
- Perham RN. Biochemistry. 1991;30:8501–8512. - PubMed
-
- Perham RN. Annu Rev Biochem. 2000;69:961–1004. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
