

Growth of the normal skull vault and its alteration in craniosynostosis: insights from human genetics and experimental studies
- PMID: 16313397
- PMCID: PMC1571561
- DOI: 10.1111/j.1469-7580.2005.00475.x
Growth of the normal skull vault and its alteration in craniosynostosis: insights from human genetics and experimental studies
Abstract
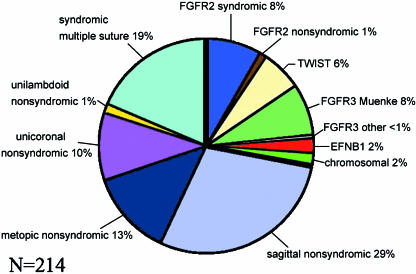
The mammalian skull vault is constructed principally from five bones: the paired frontals and parietals, and the unpaired interparietal. These bones abut at sutures, where most growth of the skull vault takes place. Sutural growth involves maintenance of a population of proliferating osteoprogenitor cells which differentiate into bone matrix-secreting osteoblasts. Sustained function of the sutures as growth centres is essential for continuous expansion of the skull vault to accommodate the growing brain. Craniosynostosis, the premature fusion of the cranial sutures, occurs in 1 in 2500 children and often presents challenging clinical problems. Until a dozen years ago, little was known about the causes of craniosynostosis but the discovery of mutations in the MSX2, FGFR1, FGFR2, FGFR3, TWIST1 and EFNB1 genes in both syndromic and non-syndromic cases has led to considerable insights into the aetiology, classification and developmental pathology of these disorders. Investigations of the biological roles of these genes in cranial development and growth have been carried out in normal and mutant mice, elucidating their individual and interdependent roles in normal sutures and in sutures undergoing synostosis. Mouse studies have also revealed a significant correspondence between the neural crest-mesoderm boundary in the early embryonic head and the position of cranial sutures, suggesting roles for tissue interaction in suture formation, including initiation of the signalling system that characterizes the functionally active suture.
Figures








References
-
- Anderson J, Burns HD, Enriquez-Harris P, Wilkie AOM, Heath JK. Apert syndrome mutations in fibroblast growth factor receptor 2 exhibit increased affinity for FGF ligand. Hum Mol Genet. 1998;7:1475–1483. - PubMed
-
- Beer HD, Vindevoghel L, Gait MJ, et al. Fibroblast growth factor (FGF) receptor 1-IIIb is a naturally occurring functional receptor for FGfs that is a naturally occurring functional receptor for FGfs that is preferentially expressed in the skin and brain. J Biol Chem. 2000;275:16091–16097. - PubMed
-
- Beiman M, Shilo BZ, Volk T. Heartless, a Drosophila FGF receptor homolog, is essential for cell migration and establishment of several mesodermal lineages. Genes Dev. 1996;10:2993–3002. - PubMed
-
- Bellus GA, Gaudenz K, Zackai EH, et al. Identical mutations in three different fibroblast growth factor receptor genes in autosomal dominant craniosynostosis syndromes. Nature Genet. 1996;14:174–176. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
