

Human Bex2 interacts with LMO2 and regulates the transcriptional activity of a novel DNA-binding complex
- PMID: 16314316
- PMCID: PMC1298925
- DOI: 10.1093/nar/gki964
Human Bex2 interacts with LMO2 and regulates the transcriptional activity of a novel DNA-binding complex
Abstract
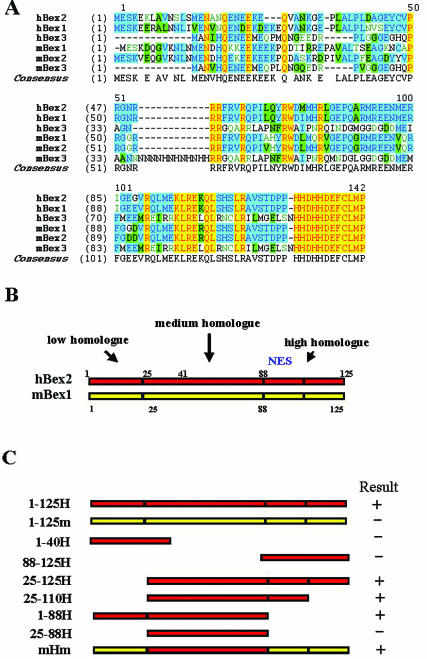
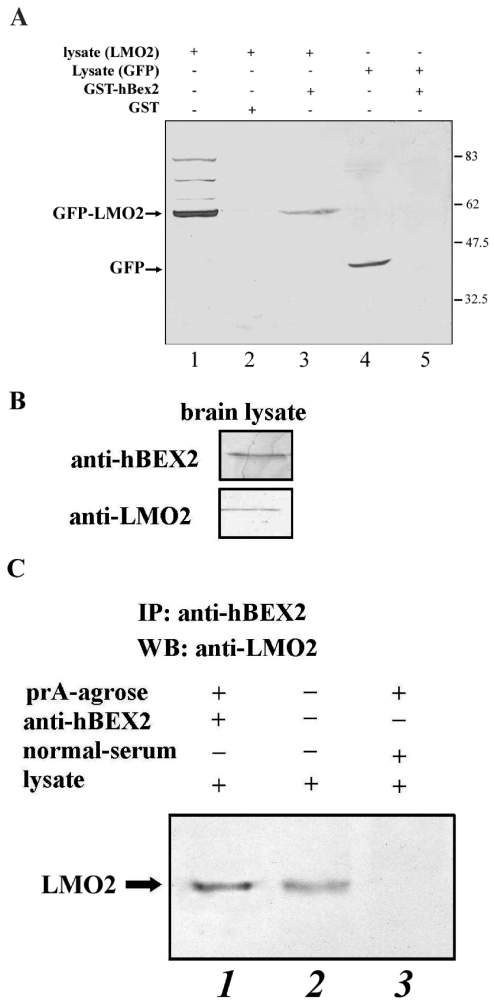
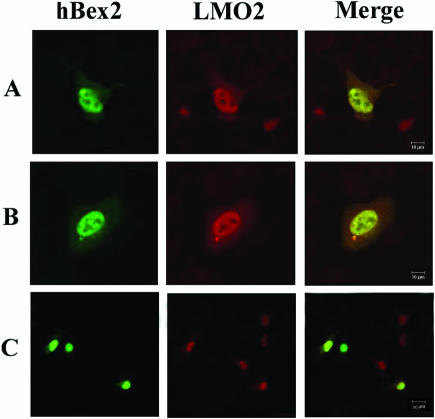
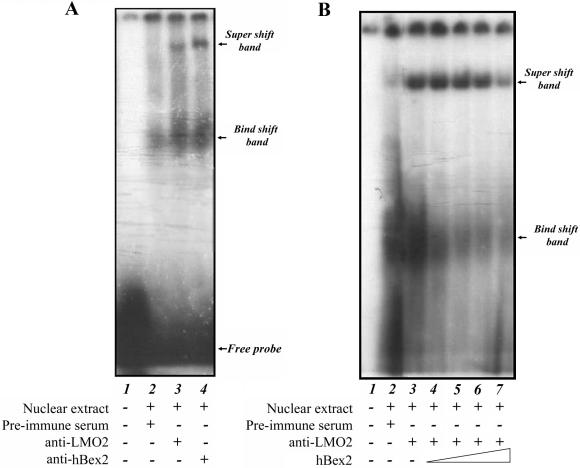
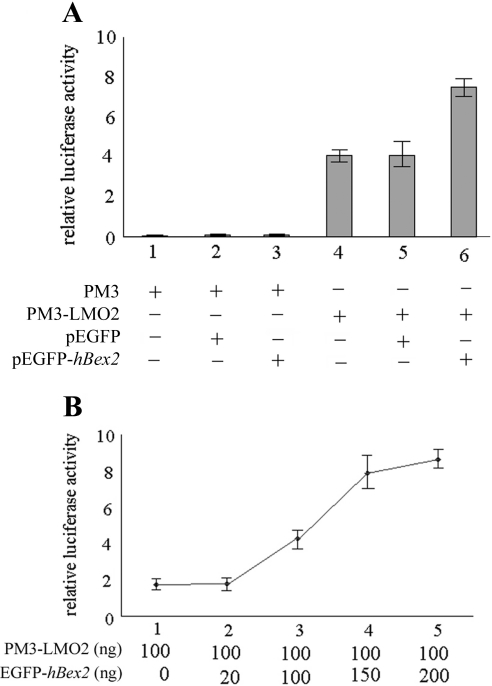
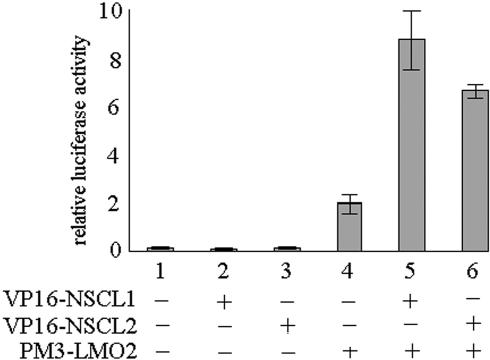
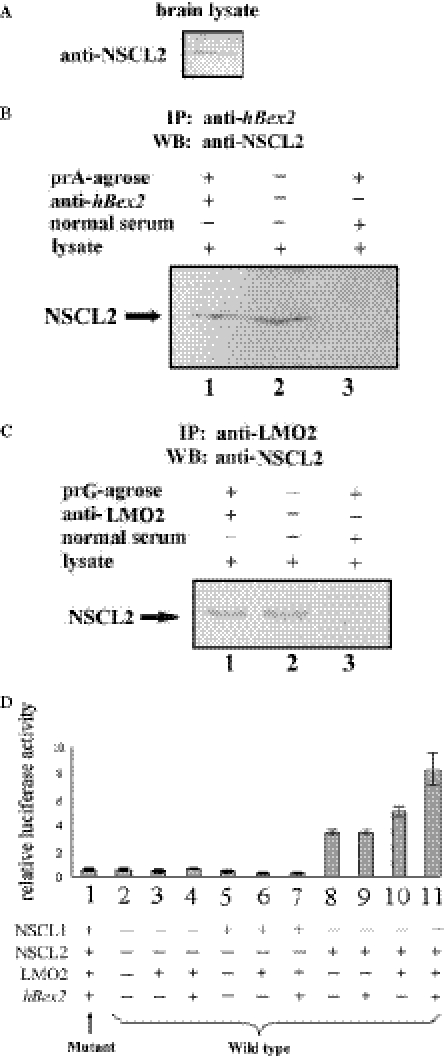
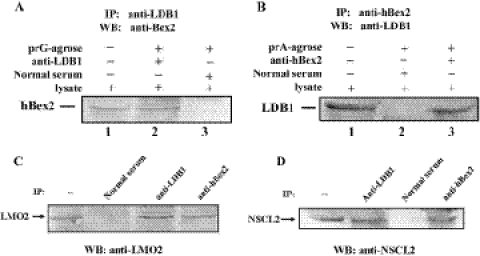
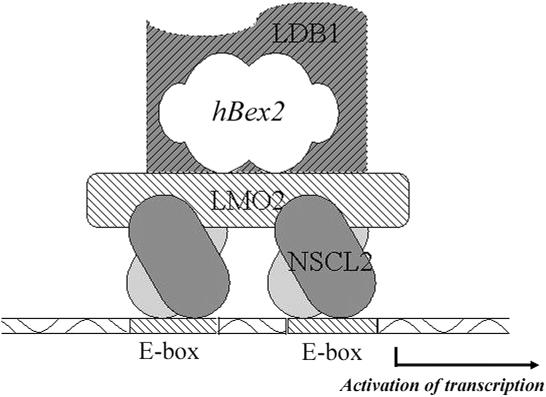
Human Bex2 (brain expressed X-linked, hBex2) is highly expressed in the embryonic brain, but its function remains unknown. We have identified that LMO2, a LIM-domain containing transcriptional factor, specifically interacts with hBex2 but not with mouse Bex1 and Bex2. The interaction was confirmed both by pull-down with GST-hBex2 and by coimmunoprecipitation assays in vivo. Using electrophoretic mobility shift assay, we have demonstrated the physical interaction of hBex2 and LMO2 as part of a DNA-binding protein complex. We have also shown that hBex2 can enhance the transcriptional activity of LMO2 in vivo. Furthermore, using mammalian two-hybrid analysis, we have identified a neuronal bHLH protein, NSCL2, as a novel binding partner for LMO2. We then showed that LMO2 could up-regulate NSCL2-dependent transcriptional activity, and hBex2 augmented this effect. Thus, hBex2 may act as a specific regulator during embryonic development by modulating the transcriptional activity of a novel E-box sequence-binding complex that contains hBex2, LMO2, NSCL2 and LDB1.
Figures
References
-
- Pan M., Yuan J., Chen B., Zhou Y., Qiang B. [Differential display analysis of human fetal brain mRNAs and isolation of brain-specific novel ESTs] Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 1997;19:93–99. - PubMed
-
- Brown A.L., Kay G.F. Bex1, a gene with increased expression in parthenogenetic embryos, is a member of a novel gene family on the mouse X chromosome. Hum. Mol. Genet. 1999;8:611–619. - PubMed
-
- Rapp G., Freudenstein J., Klaudiny J., Mucha J., Wempe F., Zimmer M., Scheit K.H. Characterization of three abundant mRNAs from human ovarian granulosa cells. DNA Cell Biol. 1990;9:479–485. - PubMed
-
- Faria T.N., LaRosa G.J., Wilen E., Liao J., Gudas L.J. Characterization of genes which exhibit reduced expression during the retinoic acid-induced differentiation of F9 teratocarcinoma cells: involvement of cyclin D3 in RA-mediated growth arrest. Mol. Cell. Endocrinol. 1998;143:155–166. - PubMed
-
- Mukai J., Hachiya T., Shoji-Hoshino S., Kimura M.T., Nadano D., Suvanto P., Hanaoka T., Li Y., Irie S., Greene L.A., et al. NADE, a p75NTR-associated cell death executor, is involved in signal transduction mediated by the common neurotrophin receptor p75NTR. J. Biol. Chem. 2000;275:17566–17570. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
