

Mutants in trs120 disrupt traffic from the early endosome to the late Golgi
- PMID: 16314430
- PMCID: PMC2171297
- DOI: 10.1083/jcb.200505145
Mutants in trs120 disrupt traffic from the early endosome to the late Golgi
Abstract
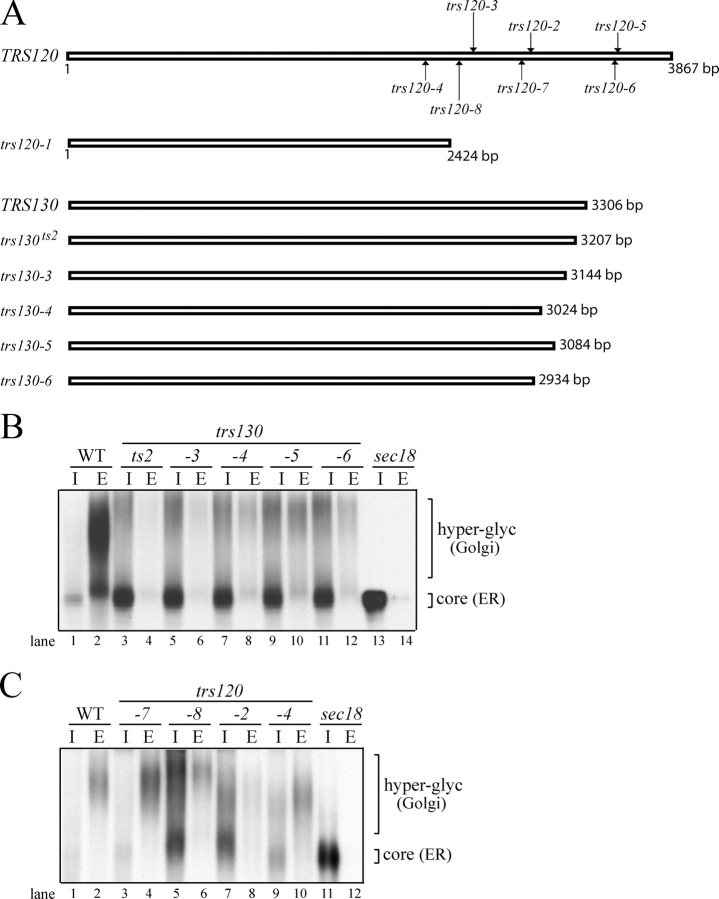
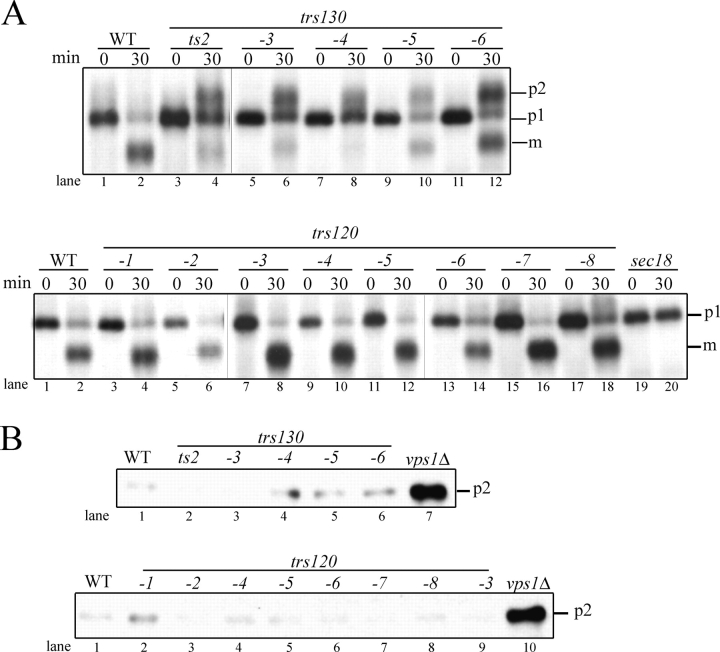
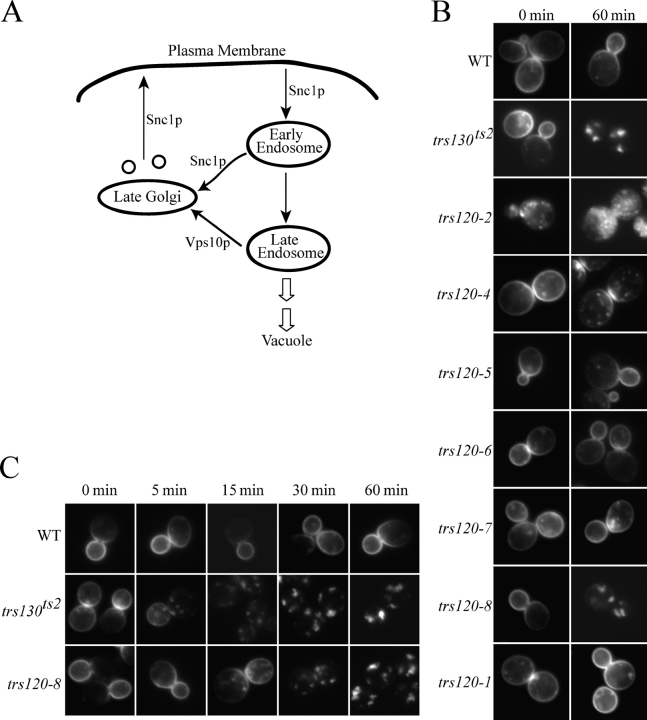
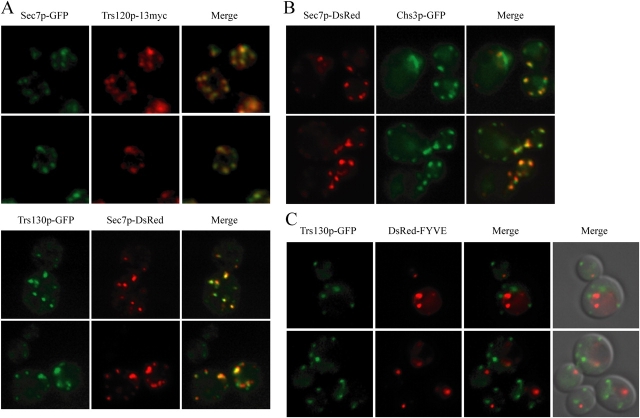
Transport protein particle (TRAPP), a large complex that mediates membrane traffic, is found in two forms (TRAPPI and -II). Both complexes share seven subunits, whereas three subunits (Trs130p, -120p, and -65p) are specific to TRAPPII. Previous studies have shown that mutations in the TRAPPII-specific gene trs130 block traffic through or from the Golgi. Surprisingly, we report that mutations in trs120 do not block general secretion. Instead, trs120 mutants accumulate aberrant membrane structures that resemble Berkeley bodies and disrupt the traffic of proteins that recycle through the early endosome. Mutants defective in recycling also display a defect in the localization of coat protein I (COPI) subunits, implying that Trs120p may participate in a COPI-dependent trafficking step on the early endosomal pathway. Furthermore, we demonstrate that Trs120p largely colocalizes with the late Golgi marker Sec7p. Our findings imply that Trs120p is required for vesicle traffic from the early endosome to the late Golgi.
Figures






References
-
- Barlowe, C., L. Orci, T. Yeung, M. Hosobuchi, S. Hamamoto, N. Salama, M.F. Rexach, M. Ravazzola, M. Amherdt, and R. Schekman. 1994. COPII: a membrane coat formed by Sec proteins that drive vesicle budding from the endoplasmic reticulum. Cell. 77:895–907. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
