

Resolution of airway inflammation and hyperreactivity after in vivo transfer of CD4+CD25+ regulatory T cells is interleukin 10 dependent
- PMID: 16314435
- PMCID: PMC1350743
- DOI: 10.1084/jem.20051166
Resolution of airway inflammation and hyperreactivity after in vivo transfer of CD4+CD25+ regulatory T cells is interleukin 10 dependent
Abstract
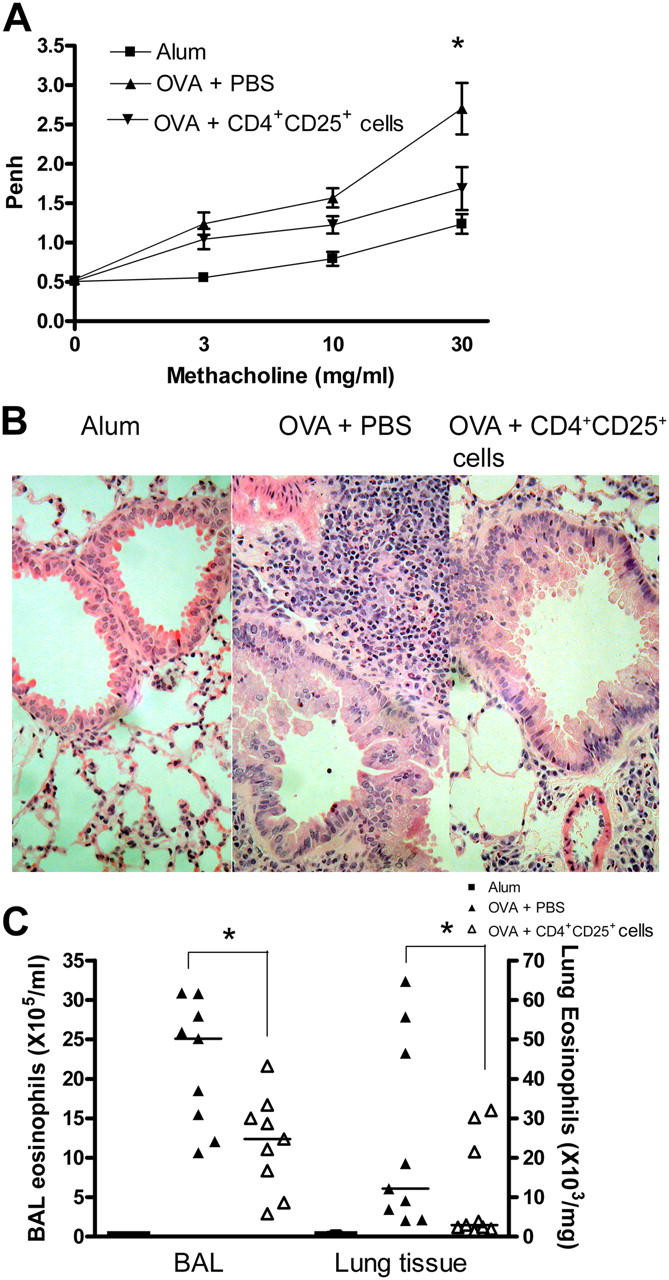
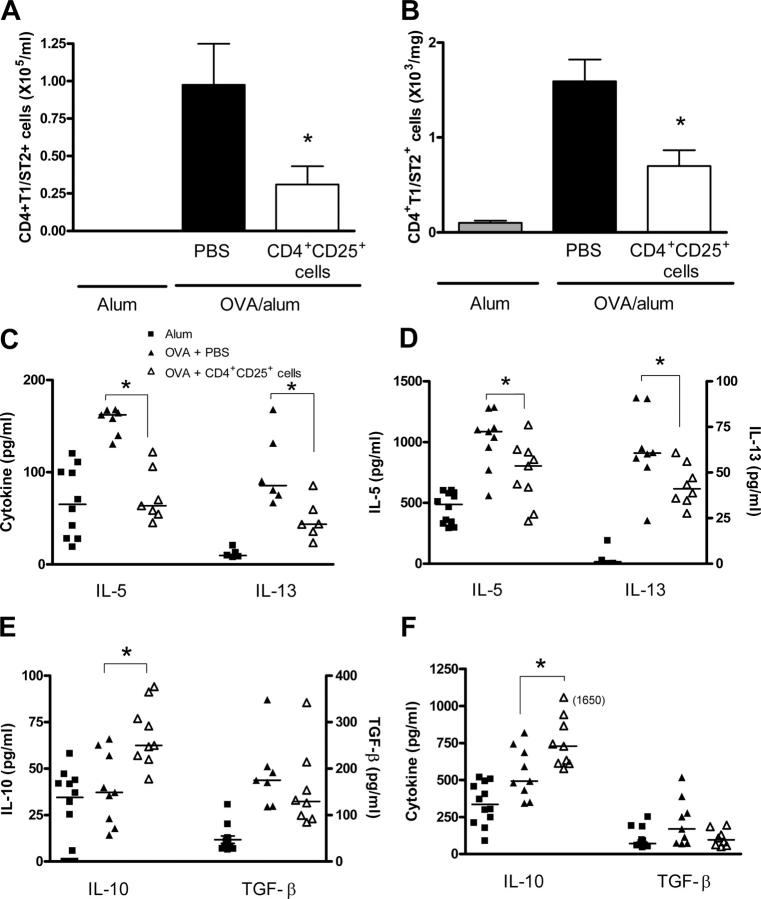
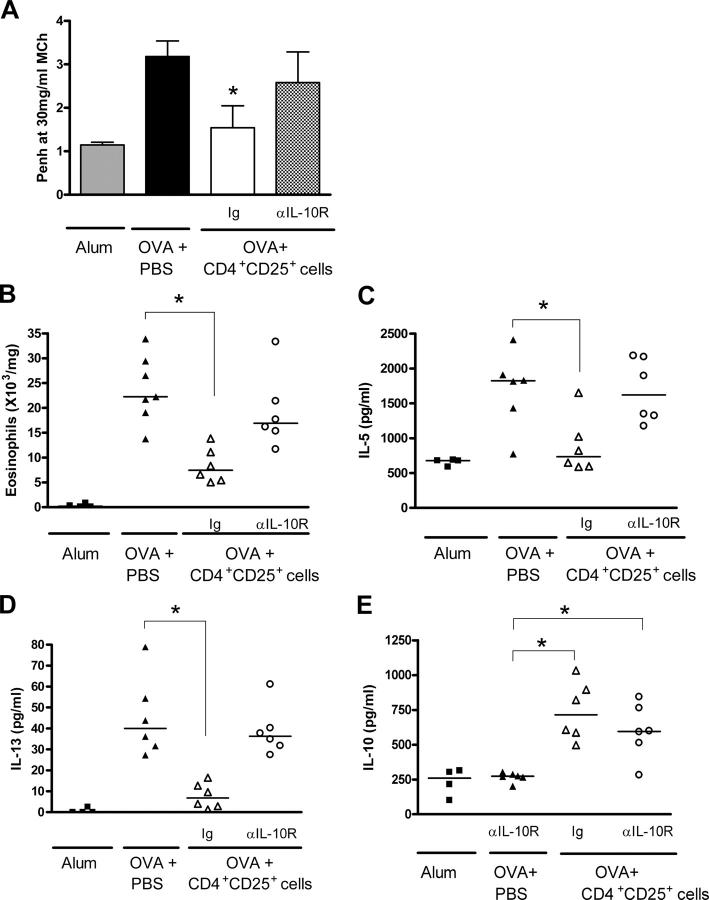
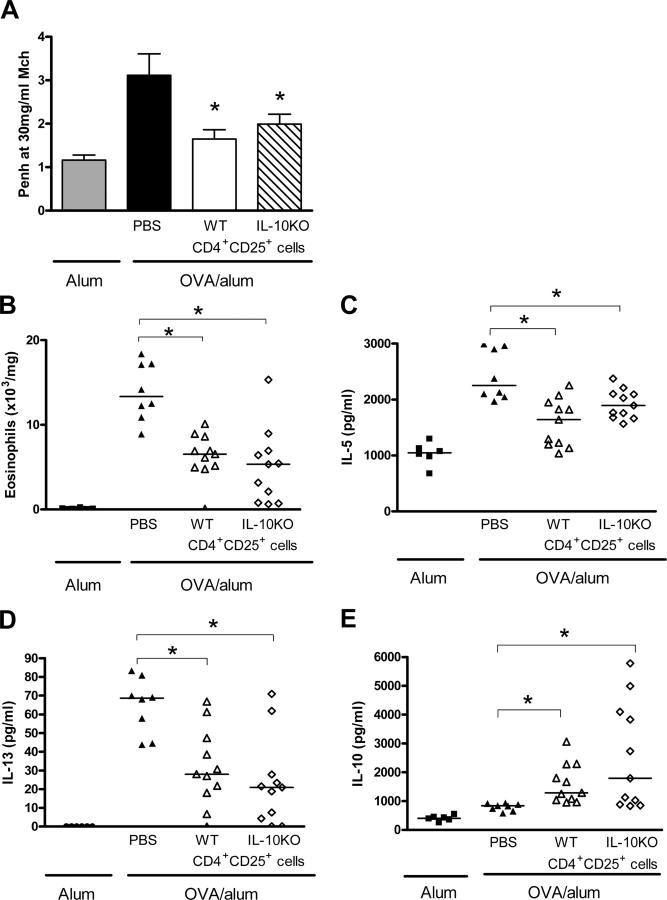
Deficient suppression of T cell responses to allergen by CD4+CD25+ regulatory T cells has been observed in patients with allergic disease. Our current experiments used a mouse model of airway inflammation to examine the suppressive activity of allergen-specific CD4+CD25+ T cells in vivo. Transfer of ovalbumin (OVA) peptide-specific CD4+CD25+ T cells to OVA-sensitized mice reduced airway hyperreactivity (AHR), recruitment of eosinophils, and T helper type 2 (Th2) cytokine expression in the lung after allergen challenge. This suppression was dependent on interleukin (IL) 10 because increased lung expression of IL-10 was detected after transfer of CD4+CD25+ T cells, and regulation was reversed by anti-IL-10R antibody. However, suppression of AHR, airway inflammation, and increased expression of IL-10 were still observed when CD4+CD25+ T cells from IL-10 gene-deficient mice were transferred. Intracellular cytokine staining confirmed that transfer of CD4+CD25+ T cells induced IL-10 expression in recipient CD4+ T cells, but no increase in IL-10 expression was detected in airway macrophages, dendritic cells, or B cells. These data suggest that CD4+CD25+ T cells can suppress the Th2 cell-driven response to allergen in vivo by an IL-10-dependent mechanism but that IL-10 production by the regulatory T cells themselves is not required for such suppression.
Figures

References
-
- Robinson, D.S., Q. Hamid, S. Ying, A. Tsicopoulos, J. Barkans, A.M. Bentley, C. Corrigan, S.R. Durham, and A.B. Kay. 1992. Predominant TH2-like bronchoalveolar T-lymphocyte population in atopic asthma. N. Engl. J. Med. 326:298–304. - PubMed
-
- Gavett, S.H., X. Chen, F. Finkelman, and M. Wills-Karp. 1994. Depletion of murine CD4+ T lymphocytes prevents antigen-induced airway hyperreactivity and pulmonary eosinophilia. Am. J. Respir. Cell Mol. Biol. 10:587–593. - PubMed
-
- Gonzalo, J.A., C.M. Lloyd, L. Kremer, E. Finger, A. Martinez, M.H. Siegelman, M. Cybulsky, and J.C. Gutierrez-Ramos. 1996. Eosinophil recruitment to the lung in a murine model of allergic inflammation. The role of T cells, chemokines, and adhesion receptors. J. Clin. Invest. 98:2332–2345. - PMC - PubMed
-
- Mosmann, T.R., and R.L. Coffman. 1989. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 7:145–173. - PubMed
-
- O'Garra, A., and P. Vieira. 2004. Regulatory T cells and mechanisms of immune system control. Nat. Med. 10:801–805. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
