

The essential WD40 protein Cia1 is involved in a late step of cytosolic and nuclear iron-sulfur protein assembly
- PMID: 16314508
- PMCID: PMC1316972
- DOI: 10.1128/MCB.25.24.10833-10841.2005
The essential WD40 protein Cia1 is involved in a late step of cytosolic and nuclear iron-sulfur protein assembly
Abstract
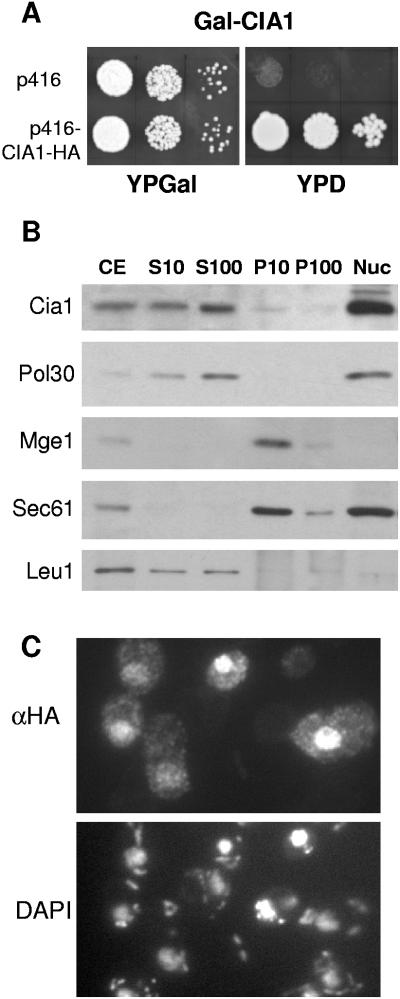
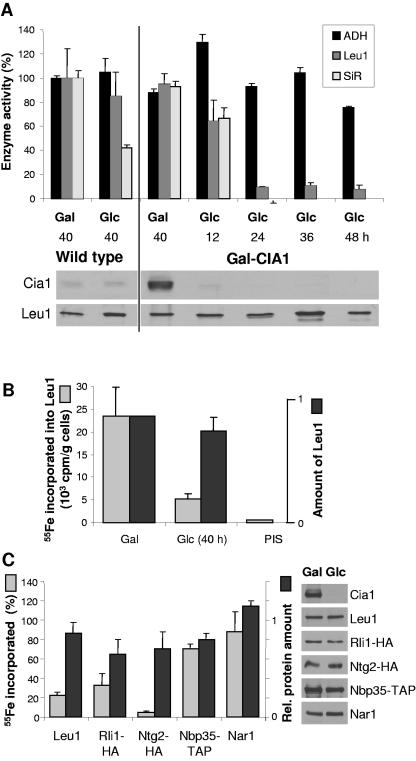
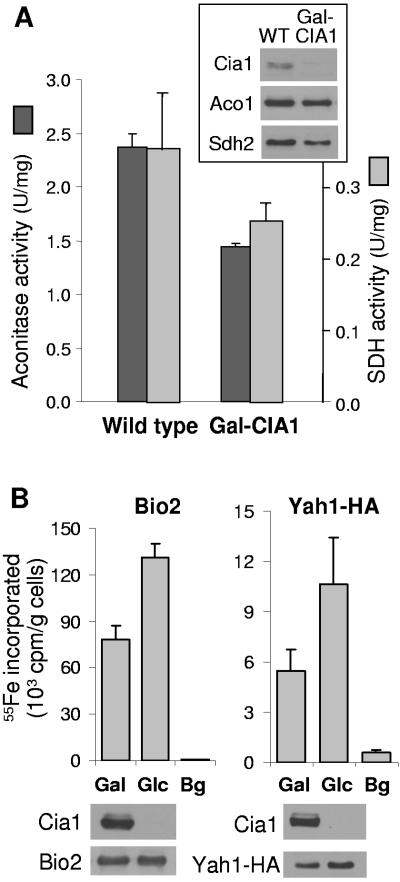
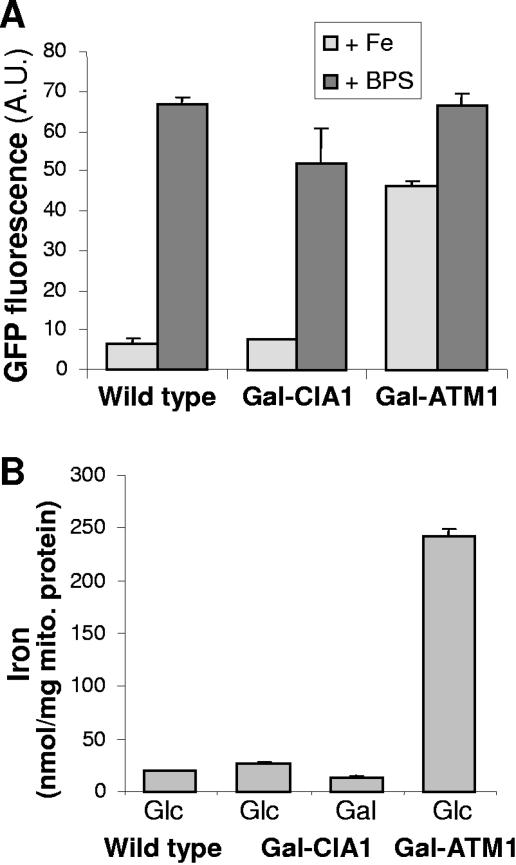
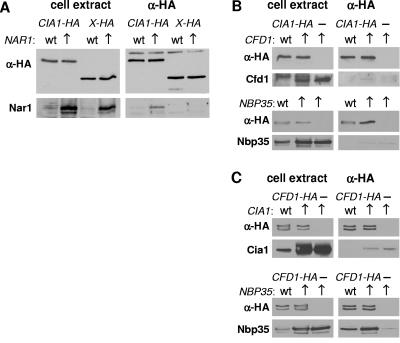
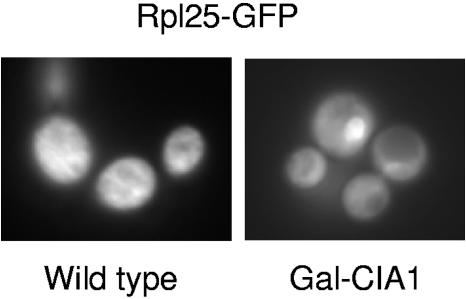
The assembly of cytosolic and nuclear iron-sulfur (Fe/S) proteins in yeast is dependent on the iron-sulfur cluster assembly and export machineries in mitochondria and three recently identified extramitochondrial proteins, the P-loop NTPases Cfd1 and Nbp35 and the hydrogenase-like Nar1. However, the molecular mechanism of Fe/S protein assembly in the cytosol is far from being understood, and more components are anticipated to take part in this process. Here, we have identified and functionally characterized a novel WD40 repeat protein, designated Cia1, as an essential component required for Fe/S cluster assembly in vivo on cytosolic and nuclear, but not mitochondrial, Fe/S proteins. Surprisingly, Nbp35 and Nar1, themselves Fe/S proteins, could assemble their Fe/S clusters in the absence of Cia1, demonstrating that these components act before Cia1. Consequently, Cia1 is involved in a late step of Fe/S cluster incorporation into target proteins. Coimmunoprecipitation assays demonstrated a specific interaction between Cia1 and Nar1. In contrast to the mostly cytosolic Nar1, Cia1 is preferentially localized to the nucleus, suggesting an additional function of Cia1. Taken together, our results indicate that Cia1 is a new member of the cytosolic Fe/S protein assembly (CIA) machinery participating in a step after Nbp35 and Nar1.
Figures
References
-
- Aris, J. P., and G. Blobel. 1991. Isolation of yeast nuclei. Methods Enzymol. 194:735-749. - PubMed
-
- Babcock, M., D. de Silva, R. Oaks, S. Davis-Kaplan, S. Jiralerspong, L. Montermini, M. Pandolfo, and J. Kaplan. 1997. Regulation of mitochondrial iron accumulation by Yfh1p, a putative homolog of frataxin. Science 276:1709-1712. - PubMed
-
- Balk, J., A. J. Pierik, D. J. Aguilar Netz, U. Mühlenhoff, and R. Lill. 2005. Nar1p, a conserved eukaryotic protein with similarity to Fe-only hydrogenases, functions in cytosolic iron-sulphur protein biogenesis. Biochem. Soc. Trans. 33:86-89. - PubMed
-
- Ghaemmaghami, S., W.-K. Huh, K. Bower, R. W. Howson, A. Belle, N. Dephoure, E. K. O'Shea, and J. S. Weissman. 2003. Global analysis of protein expression in yeast. Nature 425:737-741. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous