

Spatial periodicity of Escherichia coli K-12 biofilm microstructure initiates during a reversible, polar attachment phase of development and requires the polysaccharide adhesin PGA
- PMID: 16321928
- PMCID: PMC1317006
- DOI: 10.1128/JB.187.24.8237-8246.2005
Spatial periodicity of Escherichia coli K-12 biofilm microstructure initiates during a reversible, polar attachment phase of development and requires the polysaccharide adhesin PGA
Abstract
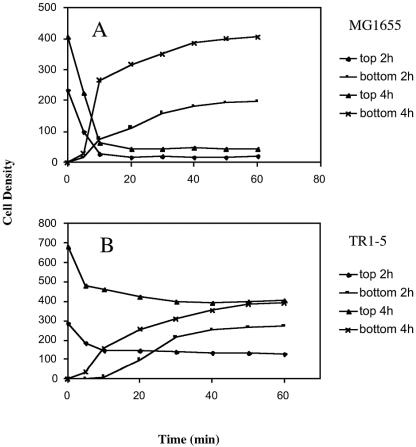
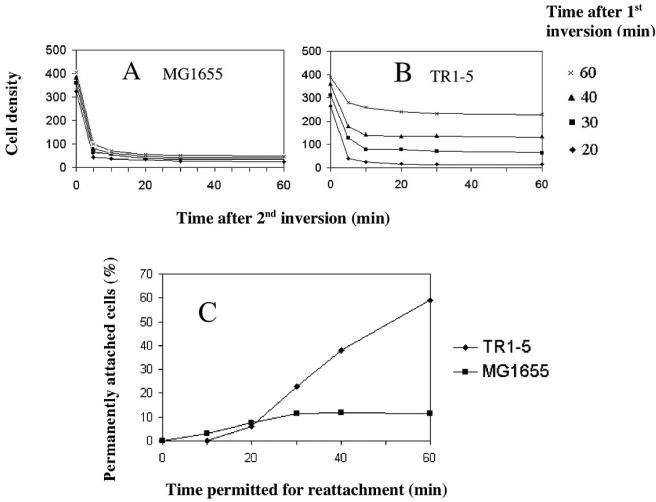
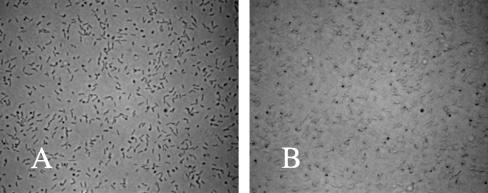
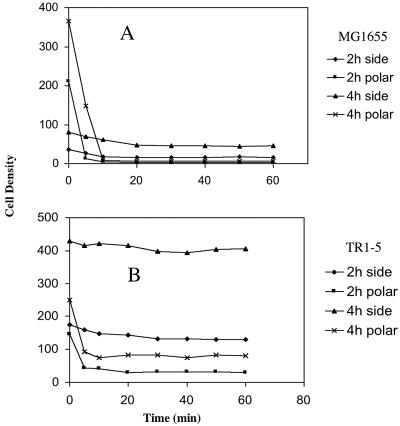
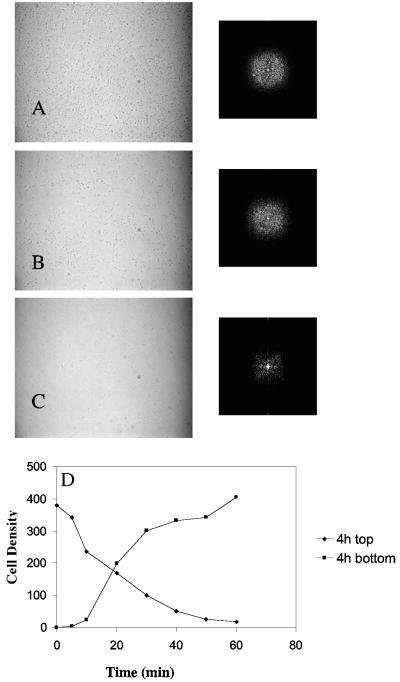
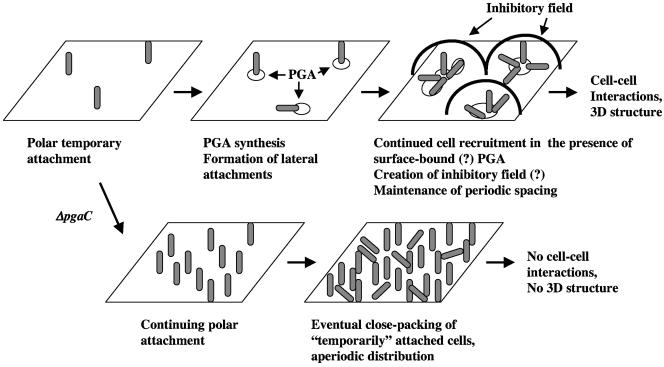
Using fast Fourier transform (FFT) analysis, we previously observed that cells within Escherichia coli biofilm are organized in nonrandom or periodic spatial patterns. Here, we developed a gravity displacement assay for examining cell adherence and used it to quantitatively monitor the formation of two distinct forms of cell attachment, temporary and permanent, during early biofilm development. Temporarily attached cells were mainly surface associated by a cell pole; permanent attachments were via the lateral cell surface. While temporary attachment precedes permanent attachment, both forms can coexist in a population. Exposure of attached cells to gravity liberated an unattached population capable of rapidly reassembling a new monolayer, composed of temporarily attached cells, and possessing periodicity. A csrA mutant, which forms biofilm more vigorously than its wild-type parent, exhibited an increased proportion of permanently attached cells and a form of attachment that was not apparent in the parent strain, permanent polar attachment. Nevertheless, it formed periodic attachment patterns. In contrast, biofilm mutants with altered lipopolysaccharide synthesis (waaG) exhibited increased cell-cell interactions, bypassed the polar attachment step, and produced FFT spectra characteristic of aperiodic cell distribution. Mutants lacking the polysaccharide adhesin beta-1,6-N-acetyl-d-glucosamine (DeltapgaC) also exhibited aperiodic cell distribution, but without apparent cell-cell interactions, and were defective in forming permanent attachments. Thus, spatial periodicity of biofilm microstructure is genetically determined and evident during the formation of temporary cell surface attachments.
Figures



References
-
- Annachhatre, A. P., and S. M. Bhamidimarri. 1992. Microbial attachment and growth in fixed-film reactors: process startup considerations. Biotechnol. Adv. 10:69-91. - PubMed
-
- Chang, I., E. S. Gilbert, N. Eliashberg, and J. D. Kiesling. 2004. A three-dimensional, stochastic simulation of biofilm growth and transport-related factors that affect structure. Microbiology 149:2859-2871. - PubMed
-
- Costerton, J. W., Z. Lewandowski, D. E. Caldwell, D. R. Korber, and H. M. Lappin-Scott. 1995. Microbial biofilms. Annu. Rev. Microbiol. 49:711-745. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
