

Timing of induction of osmotically controlled genes in Salmonella enterica Serovar Typhimurium, determined with quantitative real-time reverse transcription-PCR
- PMID: 16332813
- PMCID: PMC1317391
- DOI: 10.1128/AEM.71.12.8273-8283.2005
Timing of induction of osmotically controlled genes in Salmonella enterica Serovar Typhimurium, determined with quantitative real-time reverse transcription-PCR
Abstract
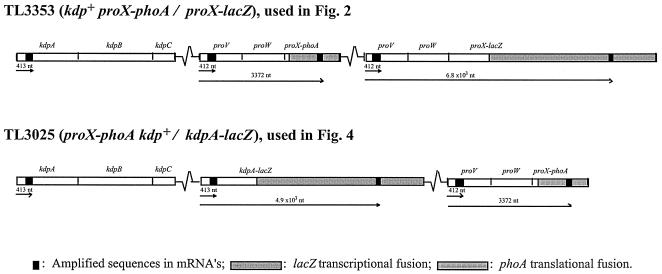
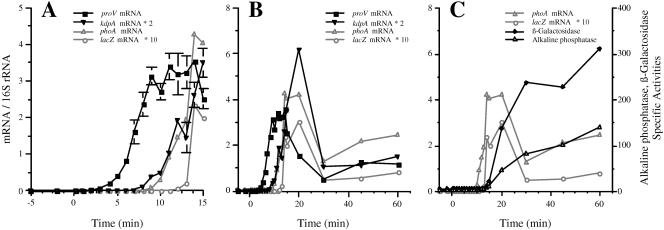
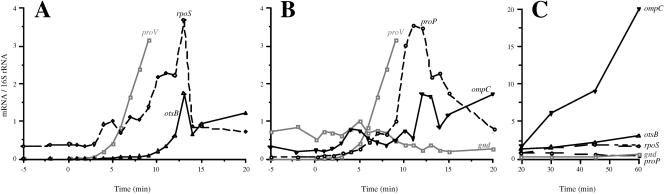
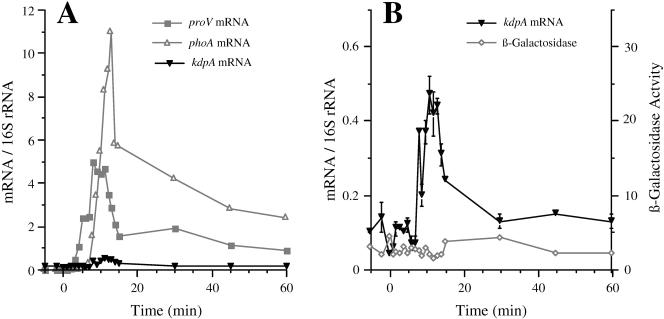
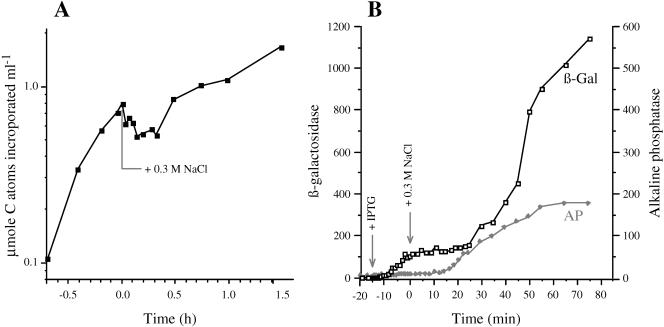
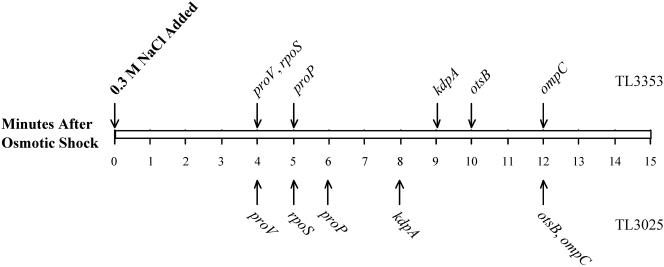
The signals that control the transcription of osmoregulated genes are not understood satisfactorily. The "turgor control model" suggested that the primary osmoregulatory signal in Enterobacteriaceae is turgor loss, which induces the kdp K+ transport operon and activates the Trk K+ permease. The ensuing increase in cytoplasmic K+ concentration was proposed to be the signal that turns on all secondary responses, including the induction of the proU (proline-glycine betaine transport) operon. The "ionic strength model" proposed that the regulatory signal for all osmotically controlled responses is the increase in the cytoplasmic ionic strength or macromolecular crowding after an osmotic upshift. The assumption in the turgor control model that the induction of kdp is a primary response to osmotic shock predicts that this response should precede all secondary responses. Both models predict that the induction of all osmotically activated responses should be independent of the chemical nature of the solute used to impose osmotic stress. We tested these predictions by quantitative real-time reverse transcription-PCR analysis of the expression of six osmotically regulated genes in Salmonella enterica serovar Typhimurium. After shock with 0.3 M NaCl, proU was induced at 4 min, proP and rpoS were induced at 4 to 6 min, kdp was induced at 8 to 9 min, and otsB and ompC were induced at 10 to 12 min. After an equivalent osmotic shock with 0.6 M sucrose, proU was induced with kinetics similar to those seen with NaCl, but induction of kdp was reduced 150-fold in comparison to induction by NaCl. Our results are inconsistent with both the turgor control and the ionic strength control models.
Figures
References
-
- Balaji, B., D. B. Bucholtz, and J. M. Anderson. 2003. Barley yellow dwarf virus and cereal yellow dwarf virus quantification by real-time polymerase chain reaction in resistant and susceptible plants. Phytopathology 93:1386-1392. - PubMed
-
- Booth, I. R., and C. F. Higgins. 1990. Enteric bacteria and osmotic stress; intracellular potassium glutamate as a secondary signal of osmotic stress. FEMS Microbiol. Rev. 75:239-246. - PubMed
-
- Bremer, H., and P. P. Dennis. 1996. Modulation of chemical composition and other parameters of the cell by growth rate, p. 1553-1569. In F. C. Neidhardt, R. Curtiss III, J. L. Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, W. S. Reznikoff, M. Riley, M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella: cellular and molecular biology, 2nd ed. American Society for Microbiology, Washington, D.C.
-
- Burg, M. B., E. D. Kwon, and D. Kültz. 1996. Osmotic regulation of gene expression. FASEB J. 10:1598-1606. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
