

Horizontal transfer of a plant transposon
- PMID: 16336045
- PMCID: PMC1310652
- DOI: 10.1371/journal.pbio.0040005
Horizontal transfer of a plant transposon
Abstract
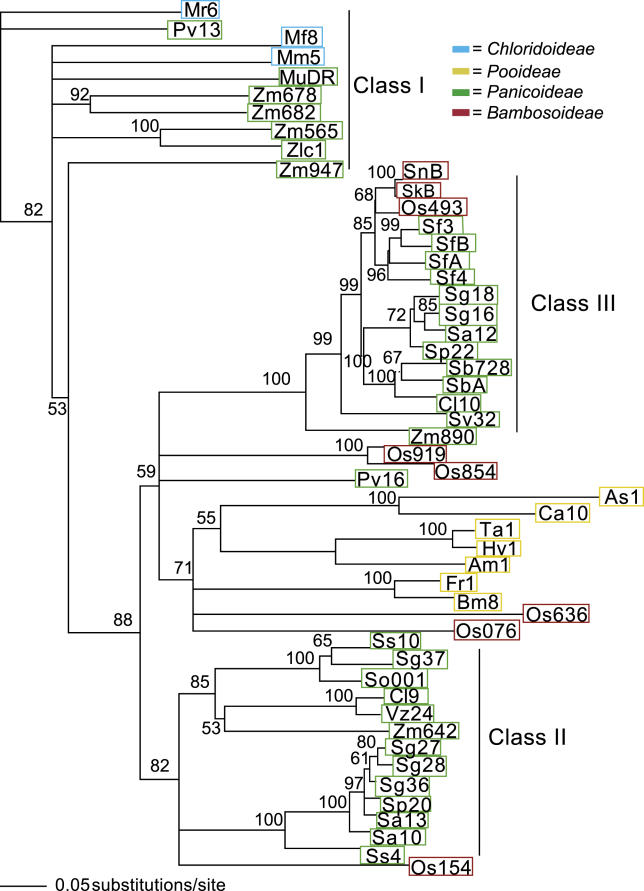
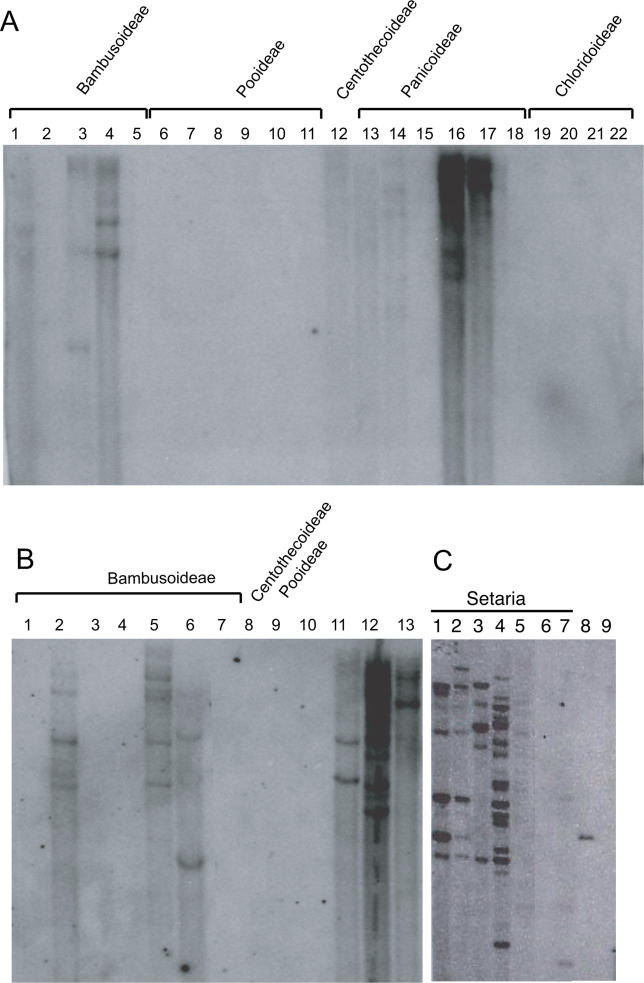
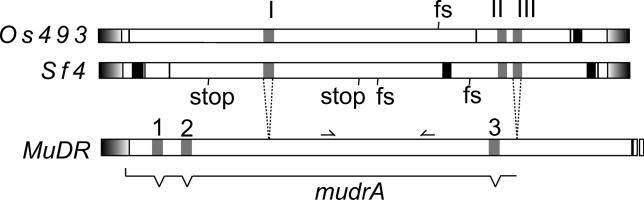
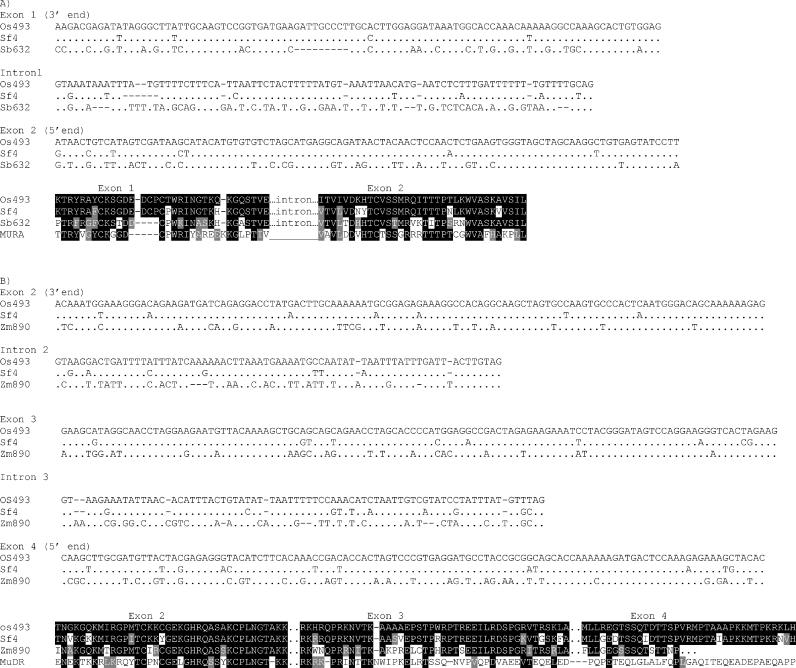
The majority of well-documented cases of horizontal transfer between higher eukaryotes involve the movement of transposable elements between animals. Surprisingly, although plant genomes often contain vast numbers of these mobile genetic elements, no evidence of horizontal transfer of a nuclear-encoded transposon between plant species has been detected to date. The most mutagenic known plant transposable element system is the Mutator system in maize. Mu-like elements (MULEs) are widespread among plants, and previous analysis has suggested that the distribution of various subgroups of MULEs is patchy, consistent with horizontal transfer. We have sequenced portions of MULE transposons from a number of species of the genus Setaria and compared them to each other and to publicly available databases. A subset of these elements is remarkably similar to a small family of MULEs in rice. A comparison of noncoding and synonymous sequences revealed that the observed similarity is not due to selection at the amino acid level. Given the amount of time separating Setaria and rice, the degree of similarity between these elements excludes the possibility of simple vertical transmission of this class of MULEs. This is the first well-documented example of horizontal transfer of any nuclear-encoded genes between higher plants.
Figures
References
-
- Jain R, Rivera MC, Moore JE, Lake JA. Horizontal gene transfer in microbial genome evolution. Theor Popul Biol. 2002;61:489–495. - PubMed
-
- Doolittle WF. Phylogenetic classification and the universal tree. Science. 1999;284:2124–2129. - PubMed
-
- Syvanen M, Kado C, editors. Horizontal gene transfer. New York: Academic Press; 2002. 445 pp.
-
- Silva JC, Kidwell MG. Horizontal transfer and selection in the evolution of P elements. Mol Biol Evol. 2000;17:1542–1557. - PubMed
-
- Robertson HM, Soto-Adames FN, Walden KO, Avancini RMP, Lampe DJ. The mariner transposons of animals: Horizontally jumping genes. In: Syvanen M, Kado CI, editors. Horizontal gene transfer. New York: Chapman and Hill; 1998. pp. 268–284.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
