

Noradrenaline triggers multivesicular release at glutamatergic synapses in the hypothalamus
- PMID: 16339033
- PMCID: PMC6725913
- DOI: 10.1523/JNEUROSCI.2378-05.2005
Noradrenaline triggers multivesicular release at glutamatergic synapses in the hypothalamus
Abstract
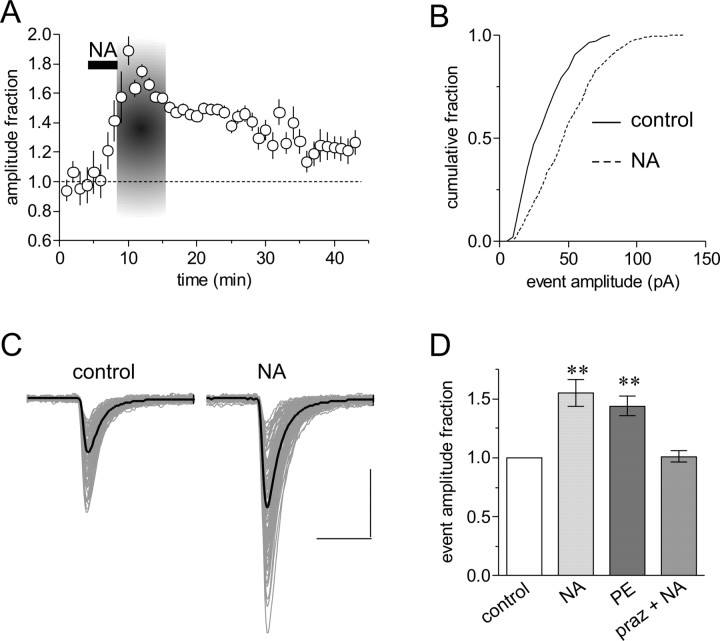
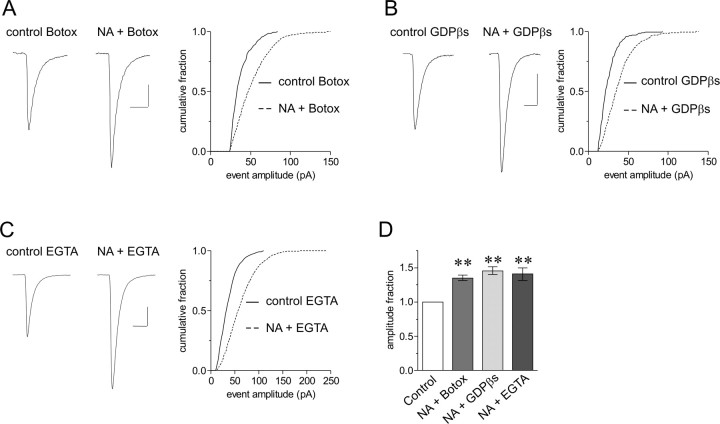
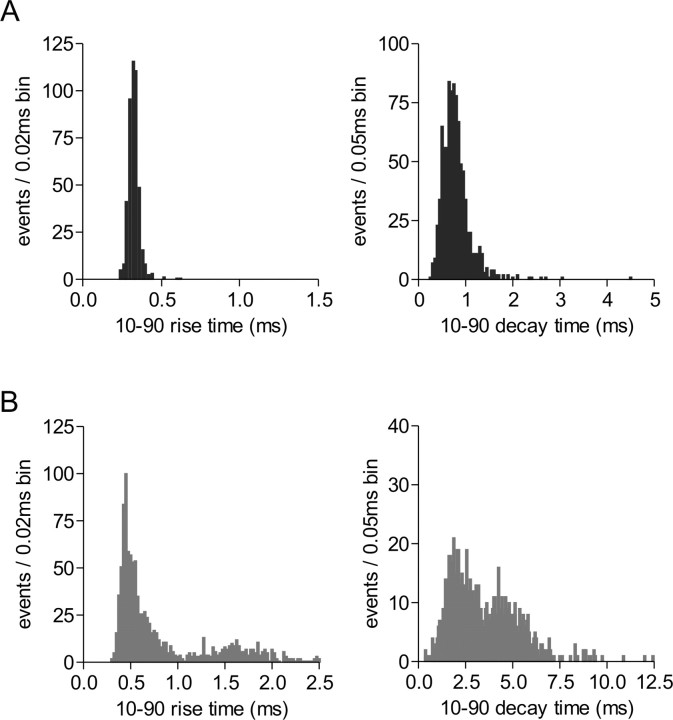
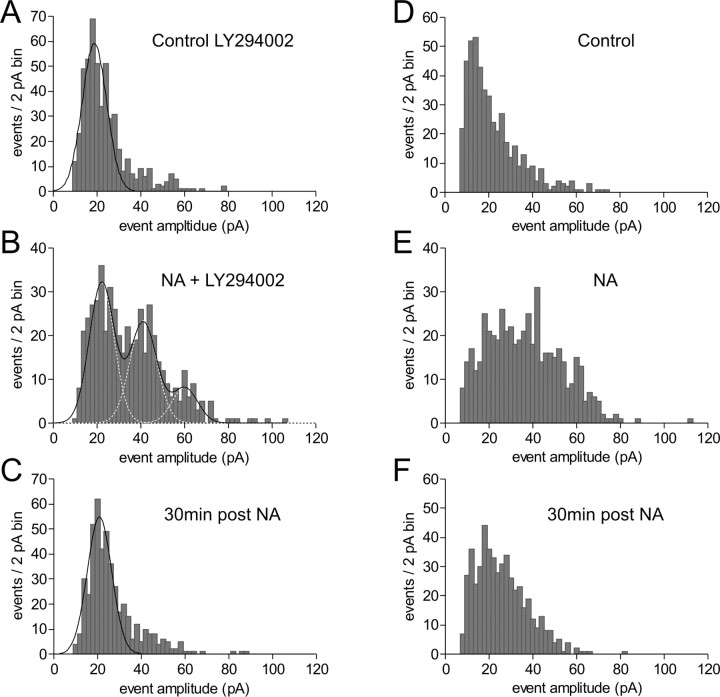
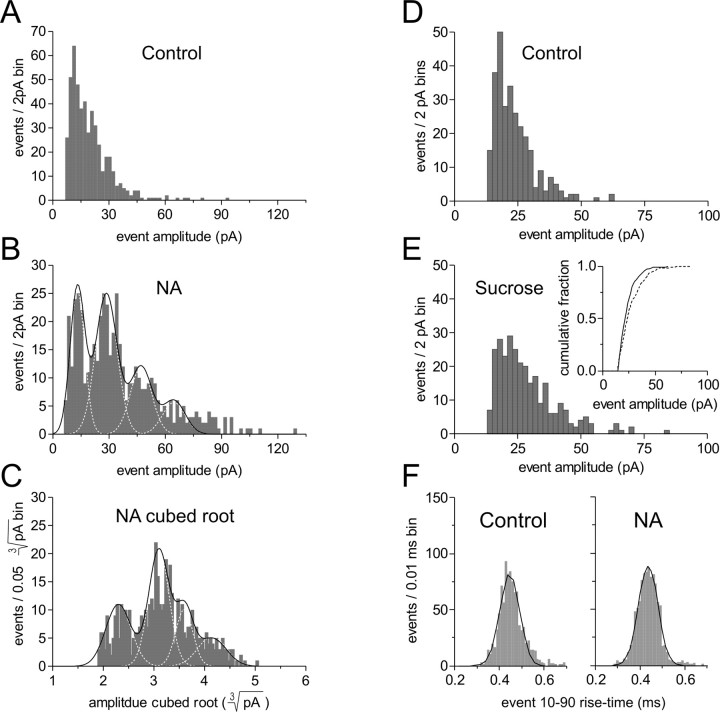
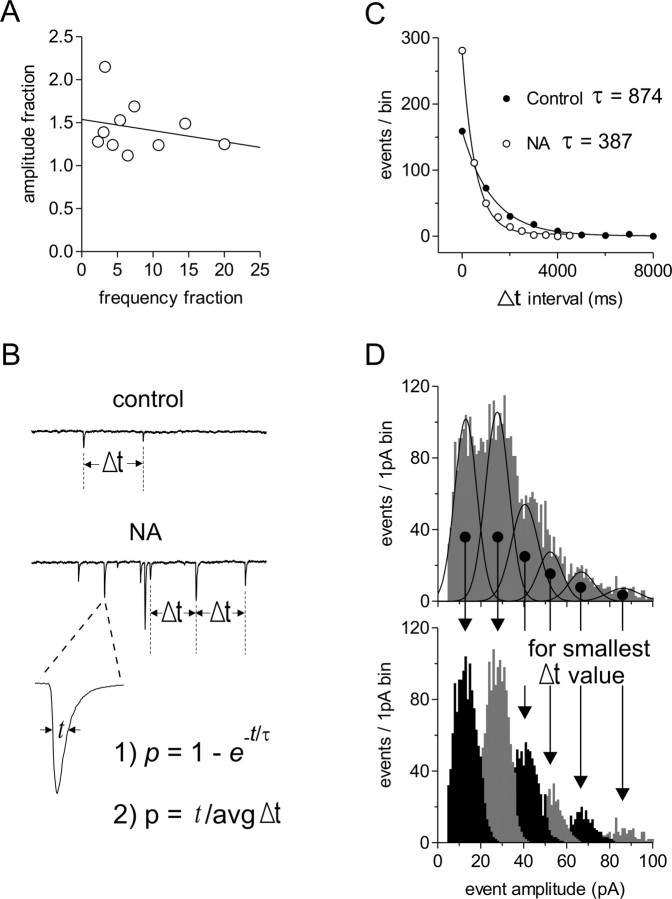
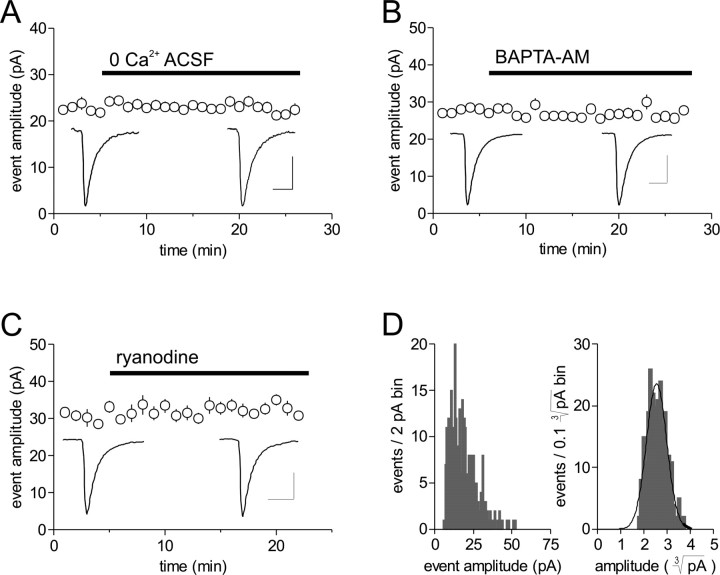
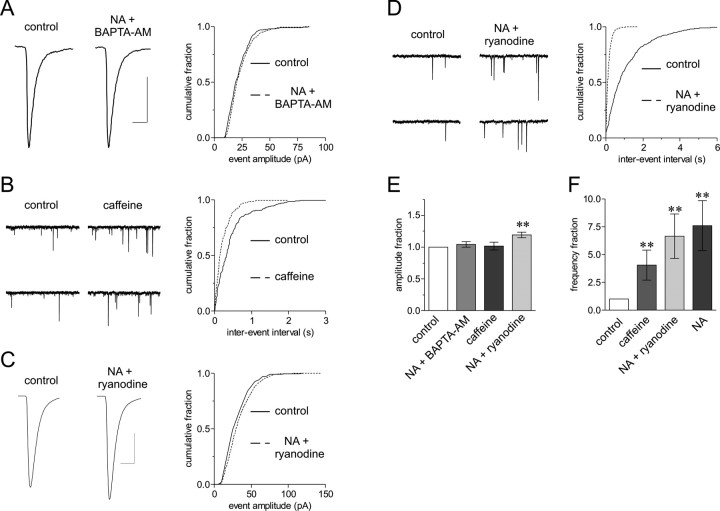
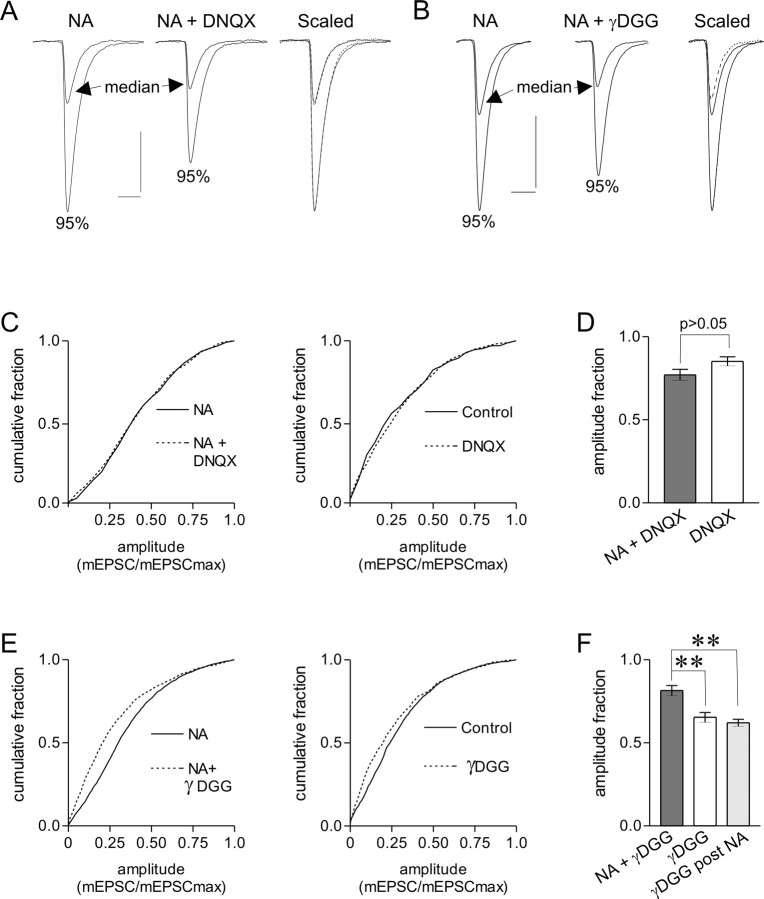
The origin of large-amplitude miniature EPSCs (mEPSCs) at central synapses remains to be firmly established. Here, we show that at excitatory synapses onto magnocellular neurosecretory cells in the hypothalamus, noradrenaline induces a rapid and robust increase in mEPSC amplitude that requires alpha1-adrenoceptor activation but is impervious to postsynaptic manipulations that block the putative insertion of AMPA receptors. In response to noradrenaline, mEPSCs exhibit a putative multimodal amplitude histogram distribution that is not attributable to random temporal summation, the unveiling of a quiescent synapse, or the release of large vesicles. Large-amplitude mEPSCs are sensitive to a high dose of ryanodine and are associated with an enhanced glutamate cleft concentration. Together, these data are consistent with the hypothesis that large-amplitude mEPSCs result from the synchronous release of multiple vesicles via rapid presynaptic calcium expulsion from intracellular stores.
Figures
References
-
- Armstrong WE, Gallagher MJ, Sladek CD (1986) Noradrenergic stimulation of supraoptic neuronal activity and vasopressin release in vitro: mediation by an alpha 1-receptor. Brain Res 365: 192-197. - PubMed
-
- Atwood HL, Karunanithi S (2002) Diversification of synaptic strength: presynaptic elements. Nat Rev Neurosci 3: 497-516. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources