

Parsing molecular and behavioral effects of cocaine in mitogen- and stress-activated protein kinase-1-deficient mice
- PMID: 16339038
- PMCID: PMC6725898
- DOI: 10.1523/JNEUROSCI.1711-05.2005
Parsing molecular and behavioral effects of cocaine in mitogen- and stress-activated protein kinase-1-deficient mice
Erratum in
- J Neurosci. 2006 Jan 18;26(3):table of contents. Simon, Arthur J [corrected to Arthur, Simon J]
Abstract
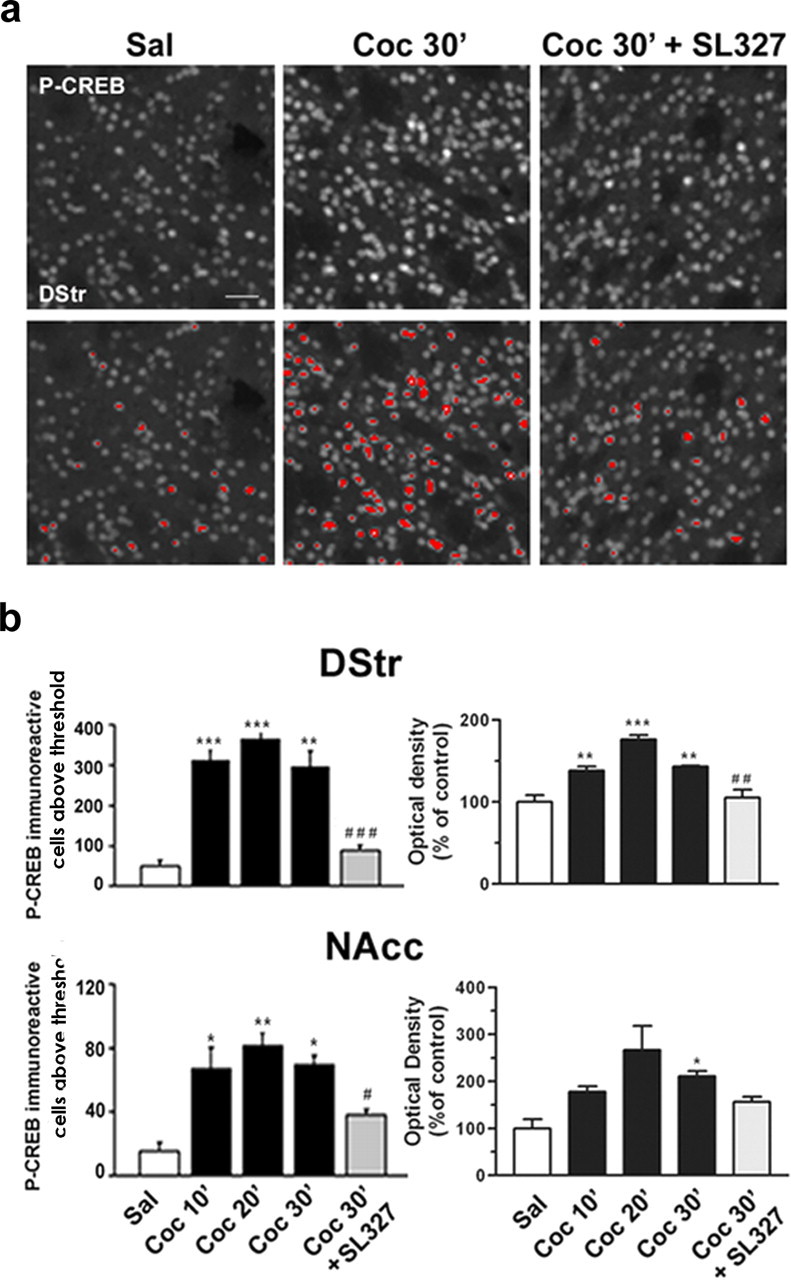
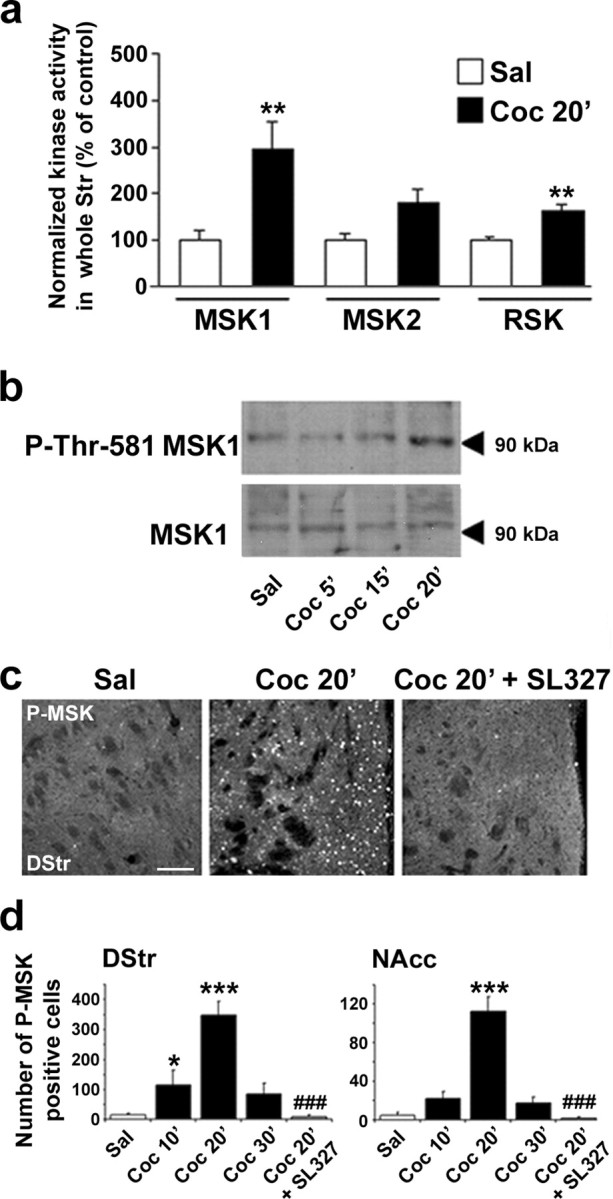
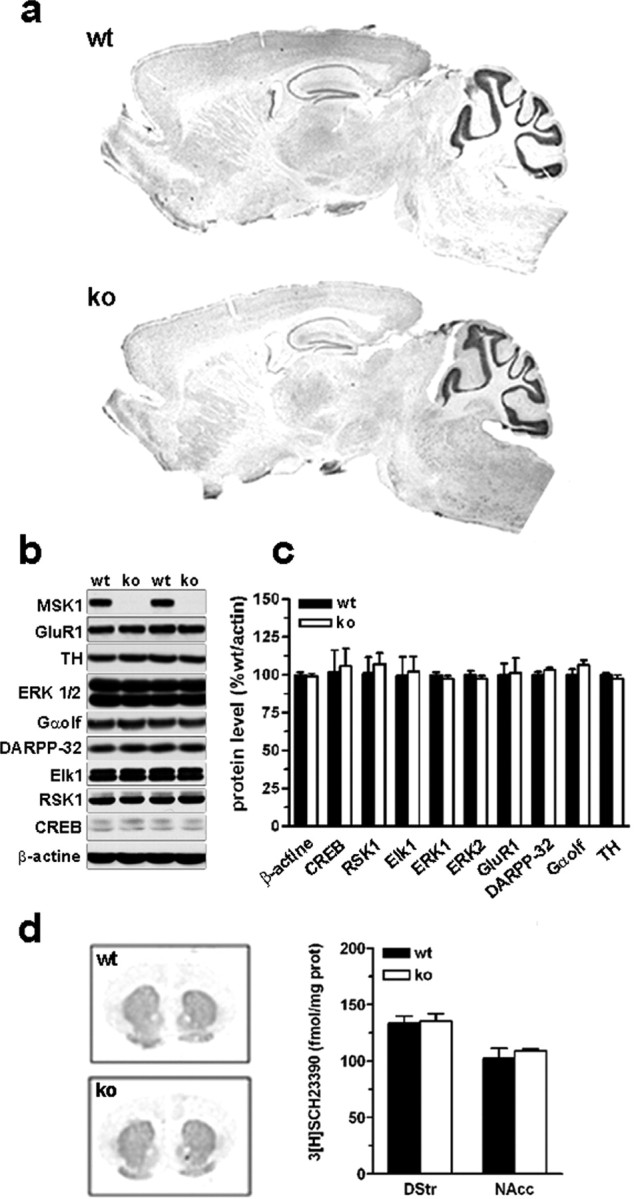
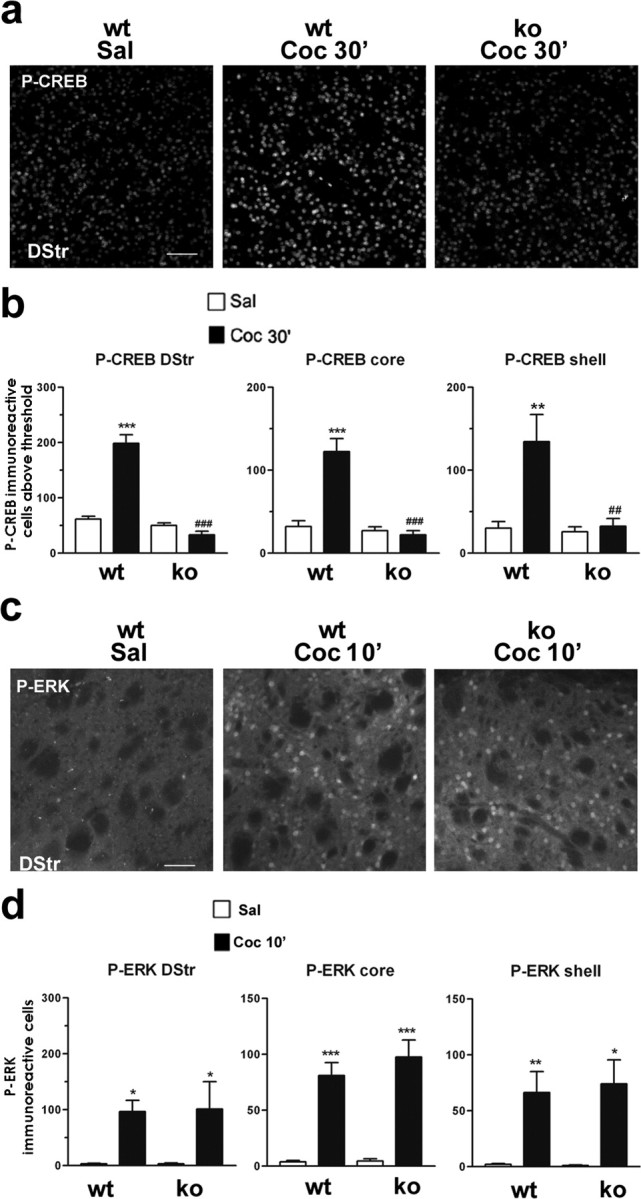
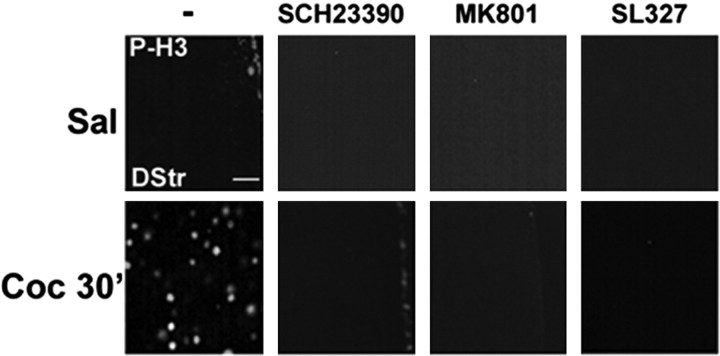
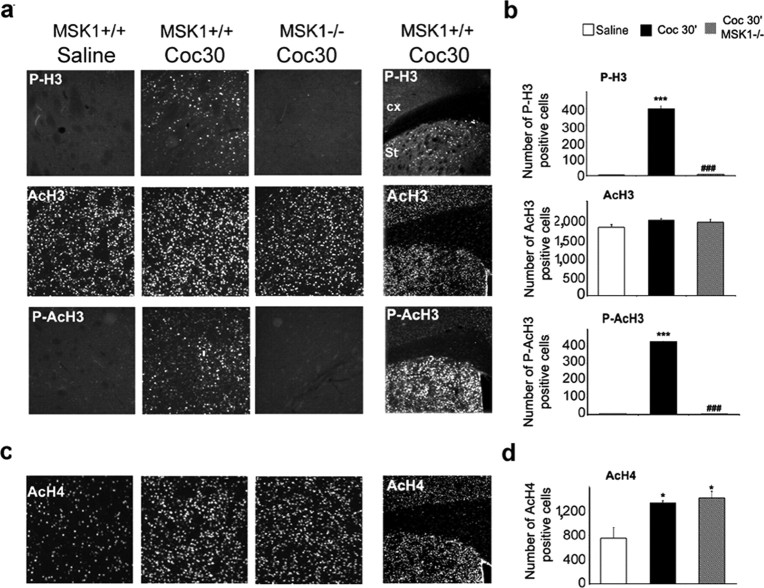
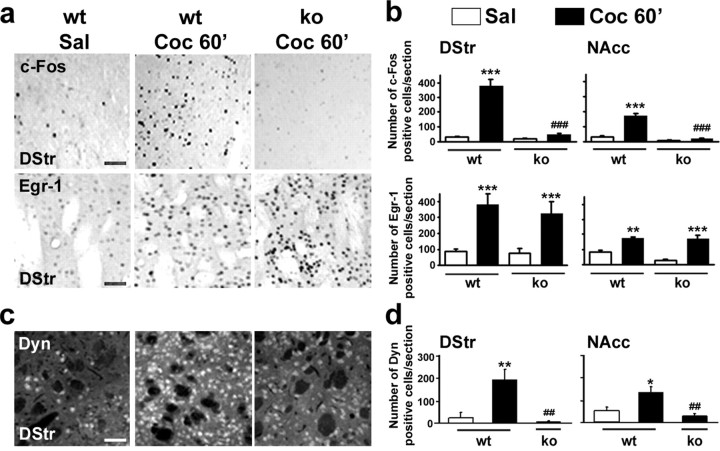
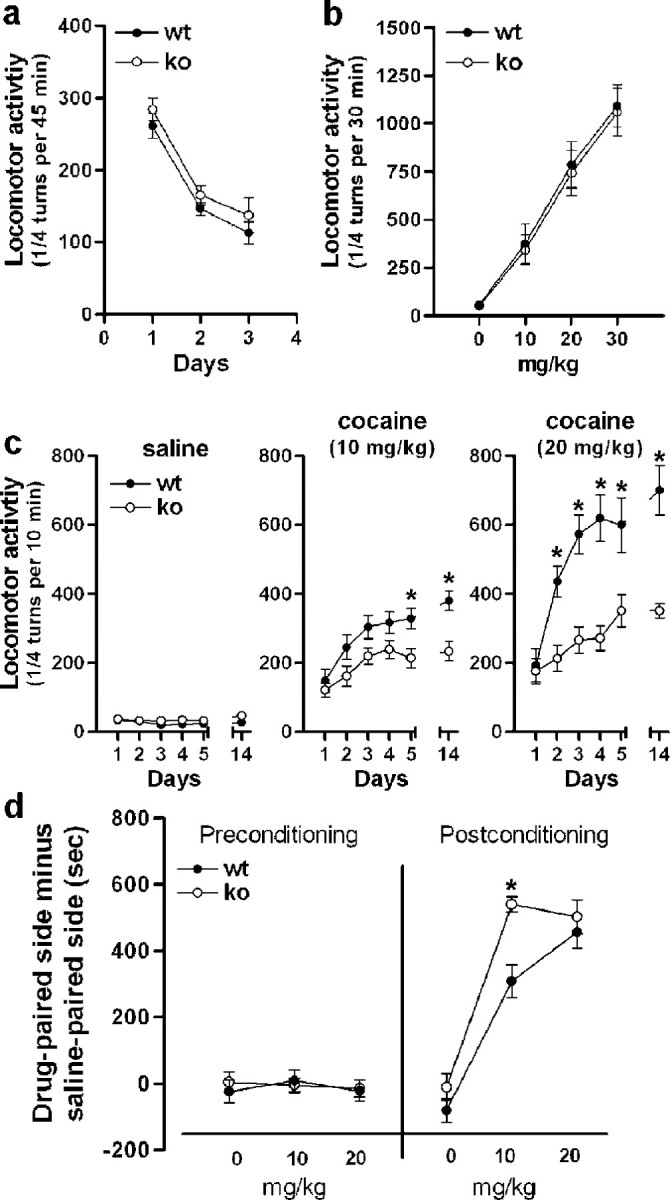
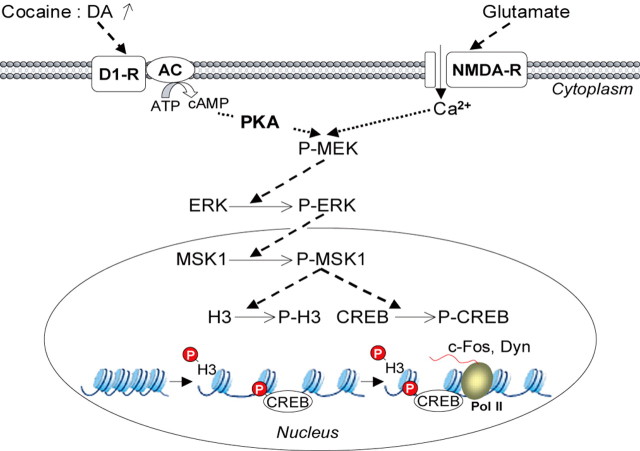
Although the induction of persistent behavioral alterations by drugs of abuse requires the regulation of gene transcription, the precise intracellular signaling pathways that are involved remain mainly unknown. Extracellular signal-regulated kinase (ERK) is critical for the expression of immediate-early genes in the striatum in response to cocaine and Delta9-tetrahydrocannabinol and for the rewarding properties of these drugs. Here we show that in mice a single injection of cocaine (10 mg/kg) activates mitogen- and stress-activated protein kinase 1 (MSK1) in dorsal striatum and nucleus accumbens. Cocaine-induced phosphorylation of MSK1 threonine 581 and cAMP response element-binding protein (CREB) serine 133 (Ser133) were blocked by SL327, a drug that prevents ERK activation. Cocaine increased the acetylation of histone H4 lysine 5 and phosphorylation of histone H3 Ser10, demonstrating the existence of drug-induced chromatin remodeling in vivo. In MSK1 knock-out (KO) mice CREB and H3 phosphorylation in response to cocaine (10 mg/kg) were blocked, and induction of c-Fos and dynorphin was prevented, whereas the induction of Egr-1 (early growth response-1)/zif268/Krox24 was unaltered. MSK1-KO mice had no obvious neurological defect but displayed a contrasted behavioral phenotype in response to cocaine. Acute effects of cocaine and dopamine D1 or D2 agonists were unaltered. Sensitivity to low doses, but not high doses, of cocaine was increased in the conditioned place preference paradigm, whereas locomotor sensitization to repeated injections of cocaine was decreased markedly. Our results show that MSK1 is a major striatal kinase, downstream from ERK, responsible for the phosphorylation of CREB and H3 and is required specifically for the induction of c-Fos and dynorphin as well as for locomotor sensitization.
Figures
References
-
- Alarcon JM, Malleret G, Touzani K, Vronskaya S, Ishii S, Kandel ER, Barco A (2004) Chromatin acetylation, memory, and LTP are impaired in CBP+/- mice: a model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration. Neuron 42: 947-959. - PubMed
-
- Arthur JS, Cohen P (2000) MSK1 is required for CREB phosphorylation in response to mitogens in mouse embryonic stem cells. FEBS Lett 482: 44-48. - PubMed
-
- Atkins CM, Selcher JC, Petraitis JJ, Trzaskos JM, Sweatt JD (1998) The MAPK cascade is required for mammalian associative learning. Nat Neurosci 1: 602-609. - PubMed
-
- Berke JD, Hyman SE (2000) Addiction, dopamine, and the molecular mechanisms of memory. Neuron 25: 515-532. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous