

SARS coronavirus, but not human coronavirus NL63, utilizes cathepsin L to infect ACE2-expressing cells
- PMID: 16339146
- PMCID: PMC8010168
- DOI: 10.1074/jbc.M508381200
SARS coronavirus, but not human coronavirus NL63, utilizes cathepsin L to infect ACE2-expressing cells
Abstract
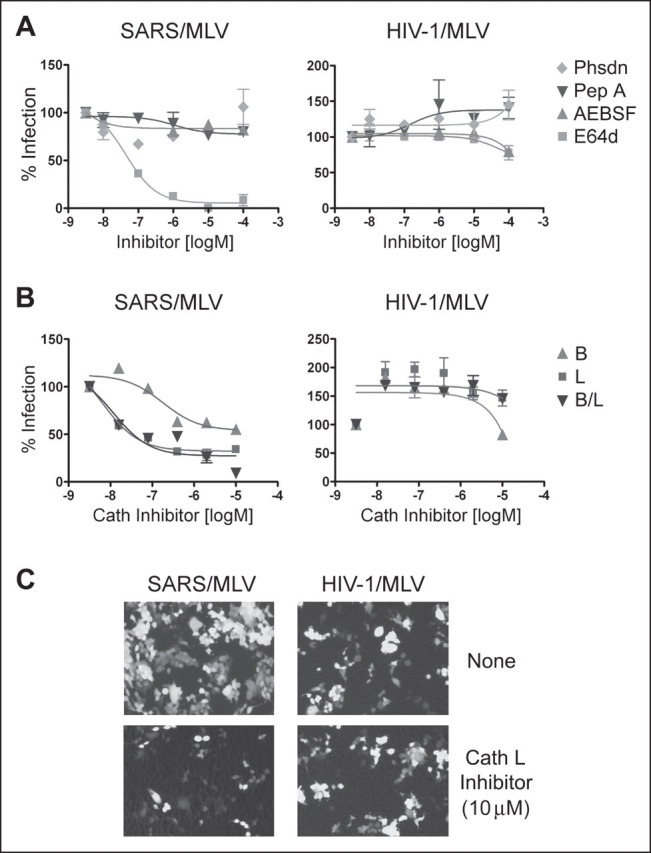
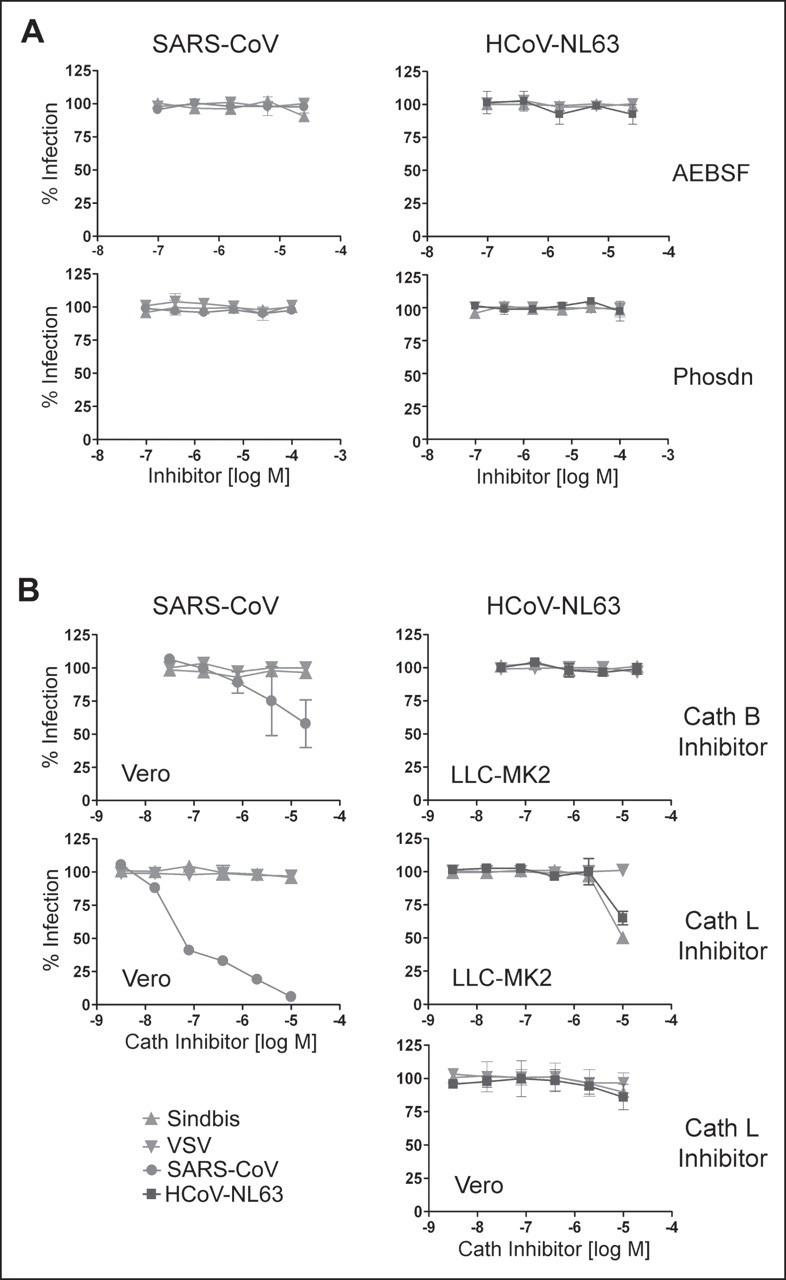
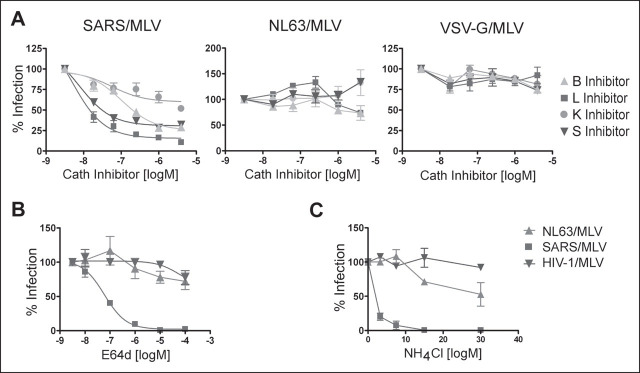
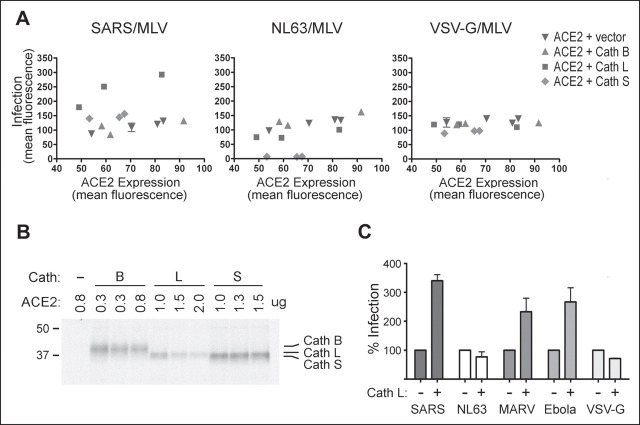
Viruses require specific cellular receptors to infect their target cells. Angiotensin-converting enzyme 2 (ACE2) is a cellular receptor for two divergent coronaviruses, SARS coronavirus (SARS-CoV) and human coronavirus NL63 (HCoV-NL63). In addition to hostcell receptors, lysosomal cysteine proteases are required for productive infection by some viruses. Here we show that SARS-CoV, but not HCoV-NL63, utilizes the enzymatic activity of the cysteine protease cathepsin L to infect ACE2-expressing cells. Inhibitors of cathepsin L blocked infection by SARS-CoV and by a retrovirus pseudotyped with the SARS-CoV spike (S) protein but not infection by HCoV-NL63 or a retrovirus pseudotyped with the HCoV-NL63 S protein. Expression of exogenous cathepsin L substantially enhanced infection mediated by the SARS-CoV S protein and by filovirus GP proteins but not by the HCoV-NL63 S protein or the vesicular stomatitis virus G protein. Finally, an inhibitor of endosomal acidification had substantially less effect on infection mediated by the HCoV-NL63 S protein than on that mediated by the SARS-CoV S protein. Our data indicate that two coronaviruses that utilize a common receptor nonetheless enter cells through distinct mechanisms.
Figures
References
-
- Kuiken T., Fouchier R.A., Schutten M., Rimmelzwaan G.F., van Amerongen G., van Riel D., Laman J.D., de Jong T., van Doornum G., Lim W., Ling A.E., Chan P.K., Tam J.S., Zambon M.C., Gopal R., Drosten C., van der Werf S., Escriou N., Manuguerra J.C., Stohr K., Peiris J.S., Osterhaus A.D. Lancet. 2003;362:263–270. - PMC - PubMed
-
- Ksiazek T.G., Erdman D., Goldsmith C.S., Zaki S.R., Peret T., Emery S., Tong S., Urbani C., Comer J.A., Lim W., Rollin P.E., Dowell S.F., Ling A.E., Humphrey C.D., Shieh W.J., Guarner J., Paddock C.D., Rota P., Fields B., DeRisi J., Yang J.Y., Cox N., Hughes J.M., LeDuc J.W., Bellini W.J., Anderson L.J. N. Engl. J. Med. 2003;348:1953–1966. - PubMed
-
- Drosten C., Gunther S., Preiser W., van der Werf S., Brodt H.R., Becker S., Rabenau H., Panning M., Kolesnikova L., Fouchier R.A., Berger A., Burguiere A.M., Cinatl J., Eickmann M., Escriou N., Grywna K., Kramme S., Manuguerra J.C., Muller S., Rickerts V., Sturmer M., Vieth S., Klenk H.D., Osterhaus A.D., Schmitz H., Doerr H.W. N. Engl. J. Med. 2003;348:1967–1976. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
