

Regulation and recovery of functions of Saccharomyces cerevisiae chaperone BiP/Kar2p after thermal insult
- PMID: 16339719
- PMCID: PMC1317487
- DOI: 10.1128/EC.4.12.2008-2016.2005
Regulation and recovery of functions of Saccharomyces cerevisiae chaperone BiP/Kar2p after thermal insult
Abstract
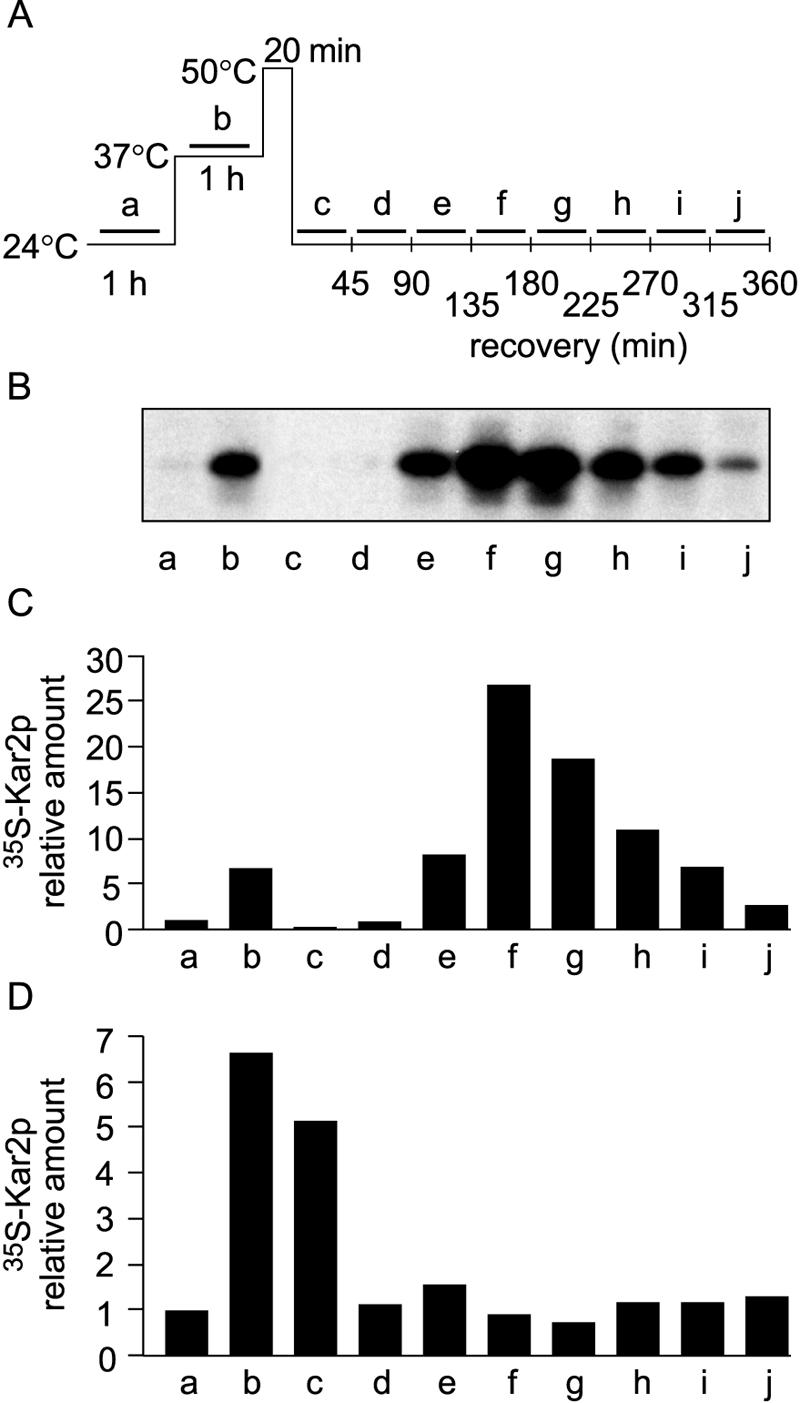
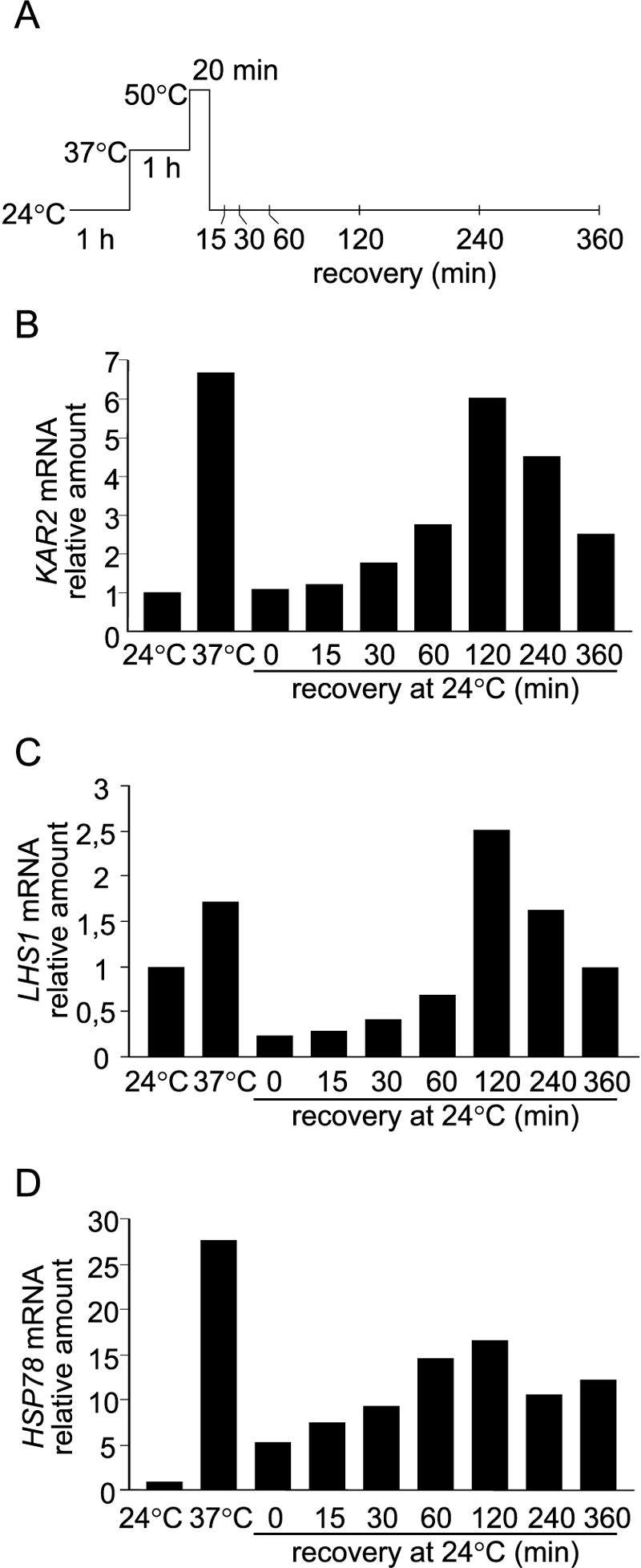
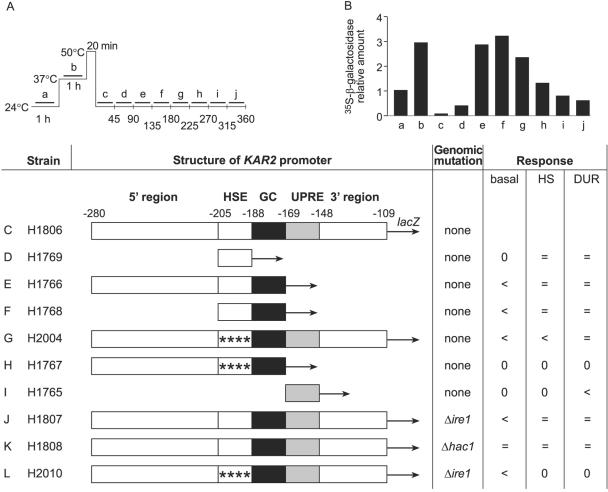
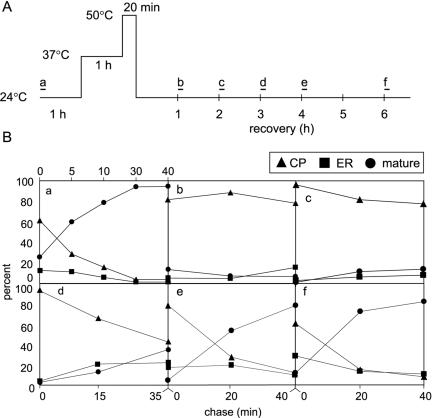
We described earlier a novel mode of regulation of Hsp104, a cytosolic chaperone directly involved in the refolding of heat-denatured proteins, and designated it delayed upregulation, or DUR. When Saccharomyces cerevisiae cells grown at the physiological temperature of 24 degrees C, preconditioned at 37 degrees C, and treated briefly at 50 degrees C were shifted back to 24 degrees C, Hsp104 expression was strongly induced after 2.5 h of recovery and returned back to normal after 5 h. Here we show that the endoplasmic reticulum (ER) chaperones BiP/Kar2p and Lhs1p and the mitochondrial chaperone Hsp78 were also upregulated at the physiological temperature during recovery from thermal insult. The heat shock element (HSE) in the KAR2 promoter was found to be sufficient to drive DUR. The unfolded protein element could also evoke DUR, albeit weakly, in the absence of a functional HSE. BiP/Kar2p functions in ER translocation and assists protein folding. Here we found that the synthesis of new BiP/Kar2p molecules was negligible for more than an hour after the shift of the cells from 50 degrees C to 24 degrees C. Concomitantly, ER translocation was blocked, suggesting that preexisting BiP/Kar2p molecules or other necessary proteins were not functioning. Translocation resumed concomitantly with enhanced synthesis of BiP/Kar2p after 3 h of recovery, after which ER exit and protein secretion also resumed. For a unicellular organism like S. cerevisiae, conformational repair of denatured proteins is the sole survival strategy. Chaperones that refold proteins in the cytosol, ER, and mitochondria of S. cerevisiae appear to be subject to DUR to ensure survival after thermal insults.
Figures
Similar articles
-
Upregulation of the Hsp104 chaperone at physiological temperature during recovery from thermal insult.Mol Microbiol. 2004 Apr;52(1):217-25. doi: 10.1111/j.1365-2958.2003.03959.x. Mol Microbiol. 2004. PMID: 15049822
-
The cytoplasmic chaperone hsp104 is required for conformational repair of heat-denatured proteins in the yeast endoplasmic reticulum.Mol Biol Cell. 1999 Nov;10(11):3623-32. doi: 10.1091/mbc.10.11.3623. Mol Biol Cell. 1999. PMID: 10564260 Free PMC article.
-
The promoter region of the yeast KAR2 (BiP) gene contains a regulatory domain that responds to the presence of unfolded proteins in the endoplasmic reticulum.Mol Cell Biol. 1993 Feb;13(2):877-90. doi: 10.1128/mcb.13.2.877-890.1993. Mol Cell Biol. 1993. PMID: 8423809 Free PMC article.
-
Role and regulation of the ER chaperone BiP.Semin Cell Dev Biol. 1999 Oct;10(5):465-72. doi: 10.1006/scdb.1999.0318. Semin Cell Dev Biol. 1999. PMID: 10597629 Review.
-
Calnexin, calreticulin, and Bip/Kar2p in protein folding.Cold Spring Harb Symp Quant Biol. 1995;60:405-15. doi: 10.1101/sqb.1995.060.01.045. Cold Spring Harb Symp Quant Biol. 1995. PMID: 8824414 Review. No abstract available.
Cited by
-
Regulation of thermotolerance by stress-induced transcription factors in Saccharomyces cerevisiae.Eukaryot Cell. 2008 May;7(5):783-90. doi: 10.1128/EC.00029-08. Epub 2008 Mar 21. Eukaryot Cell. 2008. PMID: 18359875 Free PMC article.
-
Physiological and Molecular Characterization of an Oxidative Stress-Resistant Saccharomyces cerevisiae Strain Obtained by Evolutionary Engineering.Front Microbiol. 2022 Feb 24;13:822864. doi: 10.3389/fmicb.2022.822864. eCollection 2022. Front Microbiol. 2022. PMID: 35283819 Free PMC article.
References
-
- Amorós, M., and F. Estruch. 2001. Hsf1p and Msn2/4p cooperate in the expression of Saccharomyces cerevisiae genes HSP26 and HSP104 in a gene- and stress type-dependent manner. Mol. Microbiol. 39:1523-1532. - PubMed
-
- Blond-Elguindi, S., S. E. Cwirla, W. J. Dower, R. J. Lipshutz, S. R. Sprang, J. F. Sambrook, and M. J. Gething. 1993. Affinity panning of a library of peptides displayed on bacteriophages reveals the binding specificity of BiP. Cell 75:717-728. - PubMed
-
- Caspary, F., A. Hartig, and H.-J. Schüller. 1997. Constitutive and carbon source-responsive promoter elements are involved in the regulated expression of the Saccharomyces cerevisiae malate synthase gene MLS1. Mol. Gen. Genet. 255:619-627. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
