

The polybasic juxtamembrane region of Sso1p is required for SNARE function in vivo
- PMID: 16339720
- PMCID: PMC1317504
- DOI: 10.1128/EC.4.12.2017-2028.2005
The polybasic juxtamembrane region of Sso1p is required for SNARE function in vivo
Abstract
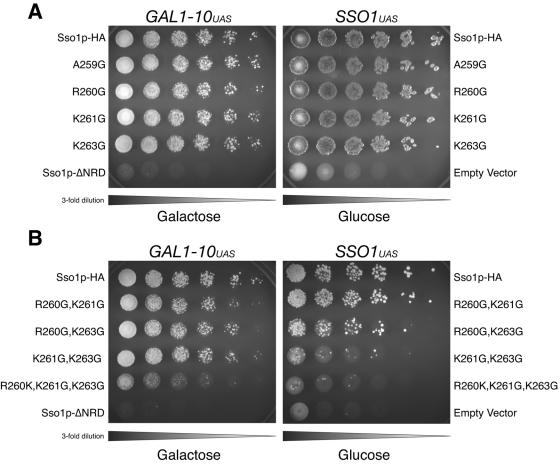
Exocytosis in Saccharomyces cerevisiae requires the specific interaction between the plasma membrane t-SNARE complex (Sso1/2p;Sec9p)and a vesicular v-SNARE (Snc1/2p). While SNARE proteins drive membrane fusion, many aspects of SNARE assembly and regulation are ill defined. Plasma membrane syntaxin homologs (including Sso1p) contain a highly charged juxtamembrane region between the transmembrane helix and the "SNARE domain" or core complex domain. We examined this region in vitro and in vivo by targeted sequence modification, including insertions and replacements. These modified Sso1 proteins were expressed as the sole copy of Sso in S. cerevisiae and examined for viability. We found that mutant Sso1 proteins with insertions or duplications show limited function, whereas replacement of as few as three amino acids preceding the transmembrane domain resulted in a nonfunctional SNARE in vivo. Viability is also maintained when two proline residues are inserted in the juxtamembrane of Sso1p, suggesting that helical continuity between the transmembrane domain and the core coiled-coil domain is not absolutely required. Analysis of these mutations in vitro utilizing a reconstituted fusion assay illustrates that the mutant Sso1 proteins are only moderately impaired in fusion. These results suggest that the sequence of the juxtamembrane region of Sso1p is vital for function in vivo, independent of the ability of these proteins to direct membrane fusion.
Figures





Similar articles
-
An intramolecular t-SNARE complex functions in vivo without the syntaxin NH2-terminal regulatory domain.J Cell Biol. 2006 Jan 16;172(2):295-307. doi: 10.1083/jcb.200507138. Epub 2006 Jan 9. J Cell Biol. 2006. PMID: 16401725 Free PMC article.
-
In vitro fusion catalyzed by the sporulation-specific t-SNARE light-chain Spo20p is stimulated by phosphatidic acid.Traffic. 2007 Nov;8(11):1630-43. doi: 10.1111/j.1600-0854.2007.00628.x. Epub 2007 Aug 14. Traffic. 2007. PMID: 17714435
-
Regulation of SNARE complex assembly by an N-terminal domain of the t-SNARE Sso1p.Nat Struct Biol. 1998 Sep;5(9):793-802. doi: 10.1038/1834. Nat Struct Biol. 1998. PMID: 9731774
-
SNARE Protein Snc1 Is Essential for Vesicle Trafficking, Membrane Fusion and Protein Secretion in Fungi.Cells. 2023 Jun 5;12(11):1547. doi: 10.3390/cells12111547. Cells. 2023. PMID: 37296667 Free PMC article. Review.
-
Syntaxin 3B: A SNARE Protein Required for Vision.Int J Mol Sci. 2024 Oct 3;25(19):10665. doi: 10.3390/ijms251910665. Int J Mol Sci. 2024. PMID: 39408994 Free PMC article. Review.
Cited by
-
Intracellular Vesicle Fusion Requires a Membrane-Destabilizing Peptide Located at the Juxtamembrane Region of the v-SNARE.Cell Rep. 2019 Dec 24;29(13):4583-4592.e3. doi: 10.1016/j.celrep.2019.11.107. Cell Rep. 2019. PMID: 31875562 Free PMC article.
-
The Dual Function of the Polybasic Juxtamembrane Region of Syntaxin 1A in Clamping Spontaneous Release and Stimulating Ca2+-Triggered Release in Neuroendocrine Cells.J Neurosci. 2018 Jan 3;38(1):220-231. doi: 10.1523/JNEUROSCI.1541-17.2017. Epub 2017 Nov 13. J Neurosci. 2018. PMID: 29133430 Free PMC article.
-
Requirement for Golgi-localized PI(4)P in fusion of COPII vesicles with Golgi compartments.Mol Biol Cell. 2011 Jan 15;22(2):216-29. doi: 10.1091/mbc.E10-04-0317. Epub 2010 Nov 30. Mol Biol Cell. 2011. PMID: 21119004 Free PMC article.
-
SNARE Proteins in Synaptic Vesicle Fusion.Adv Neurobiol. 2023;33:63-118. doi: 10.1007/978-3-031-34229-5_4. Adv Neurobiol. 2023. PMID: 37615864
-
PI(4,5)P₂-binding effector proteins for vesicle exocytosis.Biochim Biophys Acta. 2015 Jun;1851(6):785-93. doi: 10.1016/j.bbalip.2014.09.017. Epub 2014 Oct 2. Biochim Biophys Acta. 2015. PMID: 25280637 Free PMC article. Review.
References
-
- Bagnat, M., and K. Simons. 2002. Lipid rafts in protein sorting and cell polarity in budding yeast Saccharomyces cerevisiae. Biol. Chem. 383:1475-1480. - PubMed
-
- Bennett, M. K., N. Calakos, and R. H. Scheller. 1992. Syntaxin: a synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science 257:255-259. - PubMed
-
- Blumenthal, R., M. J. Clague, S. R. Durell, and R. M. Epand. 2003. Membrane fusion. Chem. Rev. 103:53-69. - PubMed
-
- Boeke, J. D., J. Trueheart, G. Natsoulis, and G. R. Fink. 1987. 5-Fluoroorotic acid as a selective agent in yeast molecular genetics. Methods Enzymol. 154:164-175. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
