

Enzymes of the heme biosynthetic pathway in the nonphotosynthetic alga Polytomella sp
- PMID: 16339726
- PMCID: PMC1317499
- DOI: 10.1128/EC.4.12.2087-2097.2005
Enzymes of the heme biosynthetic pathway in the nonphotosynthetic alga Polytomella sp
Abstract
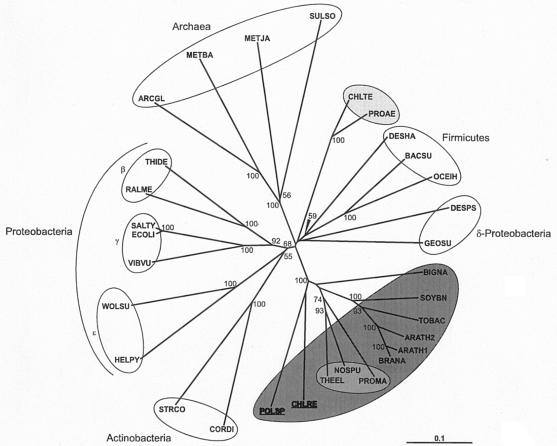
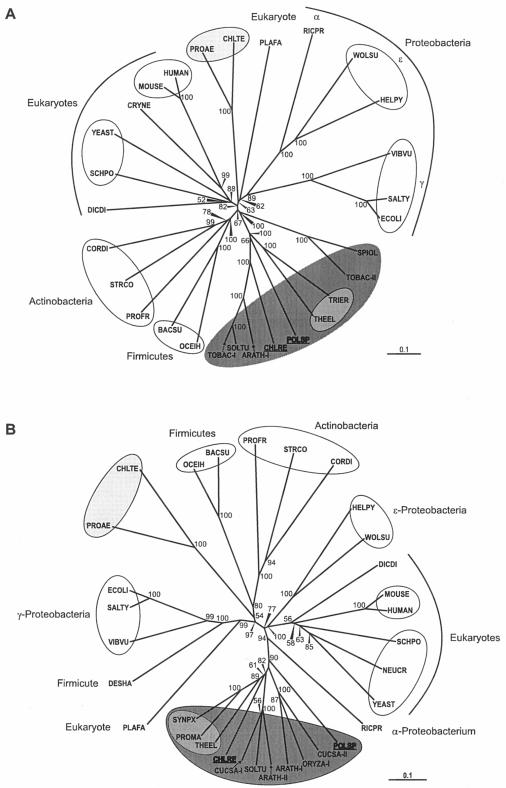
Heme biosynthesis involves a number of enzymatic steps which in eukaryotes take place in different cell compartments. Enzyme compartmentalization differs between photosynthetic and nonphotosynthetic eukaryotes. Here we investigated the structures and subcellular localizations of three enzymes involved in the heme pathway in Polytomella sp., a colorless alga evolutionarily related to the green alga Chlamydomonas reinhardtii. Functional complementation of Escherichia coli mutant strains was used to isolate cDNAs encoding three heme biosynthetic enzymes, glutamate-1-semialdehyde aminotransferase, protoporphyrinogen IX oxidase, and ferrochelatase. All three proteins show highest similarity to their counterparts in photosynthetic organisms, including C. reinhardtii. All three proteins have N-terminal extensions suggestive of intracellular targeting, and immunoblot studies indicate their enrichment in a dense cell fraction that is enriched in amyloplasts. These results suggest that even though the plastids of Polytomella sp. are not photosynthetically active, they are the major site of heme biosynthesis. The presence of a gene for glutamate-1-semialdehyde aminotransferase suggests that Polytomella sp. uses the five-carbon pathway for synthesis of the heme precursor 5-aminolevulinic acid.
Figures

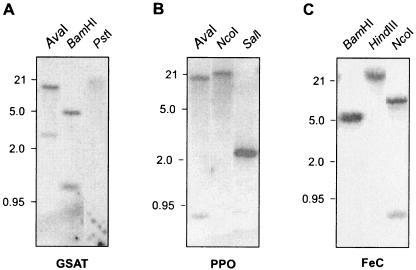
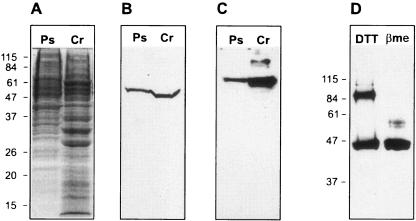
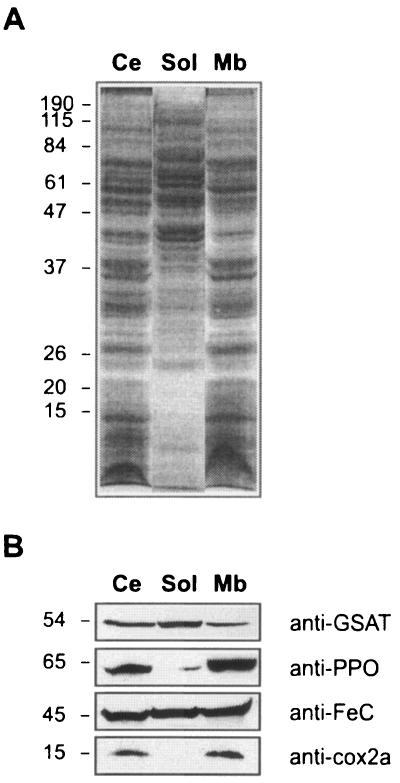
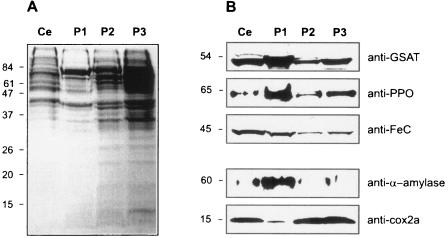
References
-
- Antaramian, A., R. Coria, J. Ramírez, and D. González-Halphen. 1996. The deduced primary structure of subunit I from cytochrome c oxidase suggests that the genus Polytomella shares a common mitochondrial origin with Chlamydomonas. Biochim. Biophys. Acta 1273:198-202. - PubMed
-
- Antaramian, A., S. Funes, M. Vásquez-Acevedo, A. Atteia, R. Coria, and D. González-Halphen. 1998. Two unusual amino acid substitutions in cytochrome b of the colorless alga Polytomella spp.: correlation with the atypical spectral properties of the bH heme. Arch. Biochem. Biophys. 354:206-214. - PubMed
-
- Atteia, A., R. van Lis, J. Ramirez, and D. González-Halphen. 2000. Polytomella spp. growth on ethanol. Extracellular pH affects the accumulation of mitochondrial cytochrome c550. Eur. J. Biochem. 267:2850-2858. - PubMed
-
- Atteia, A., R. van Lis, G. Mendoza-Hernández, K. Henze, W. Martin, H. Riveros-Rosas, and D. González-Halphen. 2003. Bifunctional aldehyde/alcohol dehydrogenase (ADHE) in chlorophyte algal mitochondria. Plant Mol. Biol. 53:175-188. - PubMed
-
- Beale, S. I. 1999. Enzymes of chlorophyll biosynthesis. Photosynth. Res. 60:43-73.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
