

Essential role of Isd11 in mitochondrial iron-sulfur cluster synthesis on Isu scaffold proteins
- PMID: 16341089
- PMCID: PMC1356349
- DOI: 10.1038/sj.emboj.7600906
Essential role of Isd11 in mitochondrial iron-sulfur cluster synthesis on Isu scaffold proteins
Abstract
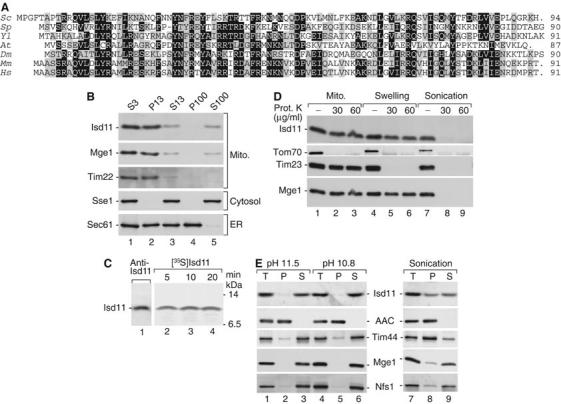
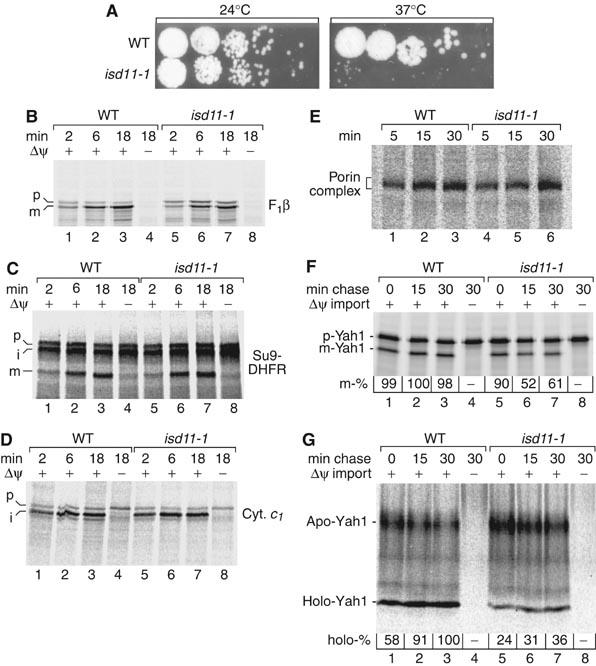
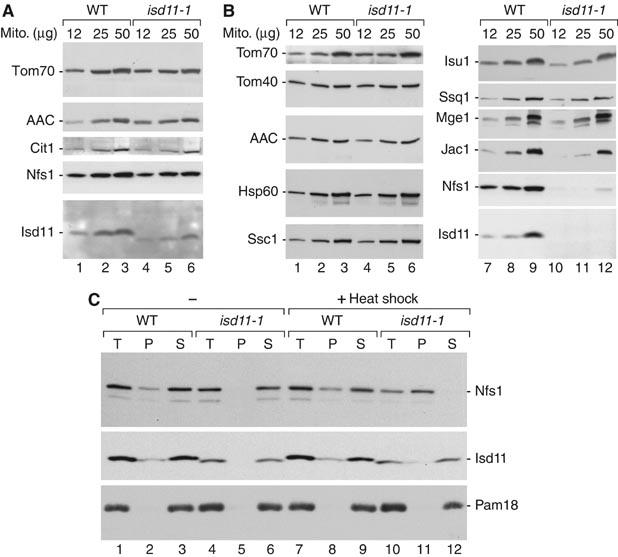
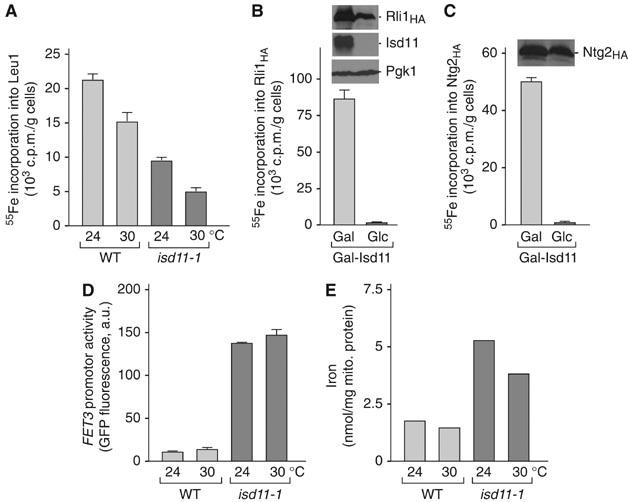
Mitochondria are indispensable for cell viability; however, major mitochondrial functions including citric acid cycle and oxidative phosphorylation are dispensable. Most known essential mitochondrial proteins are involved in preprotein import and assembly, while the only known essential biosynthetic process performed by mitochondria is the biogenesis of iron-sulfur clusters (ISC). The components of the mitochondrial ISC-assembly machinery are derived from the prokaryotic ISC-assembly machinery. We have identified an essential mitochondrial matrix protein, Isd11 (YER048w-a), that is found in eukaryotes only. Isd11 is required for biogenesis of cellular Fe/S proteins and thus is a novel subunit of the mitochondrial ISC-assembly machinery. It forms a complex with the cysteine desulfurase Nfs1 and is required for formation of an Fe/S cluster on the Isu scaffold proteins. We conclude that Isd11 is an indispensable eukaryotic component of the mitochondrial machinery for biogenesis of Fe/S proteins.
Figures

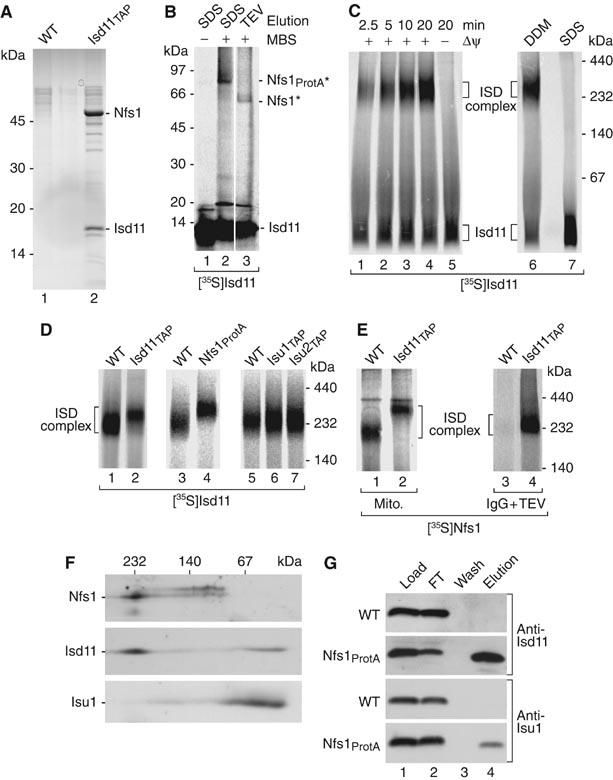
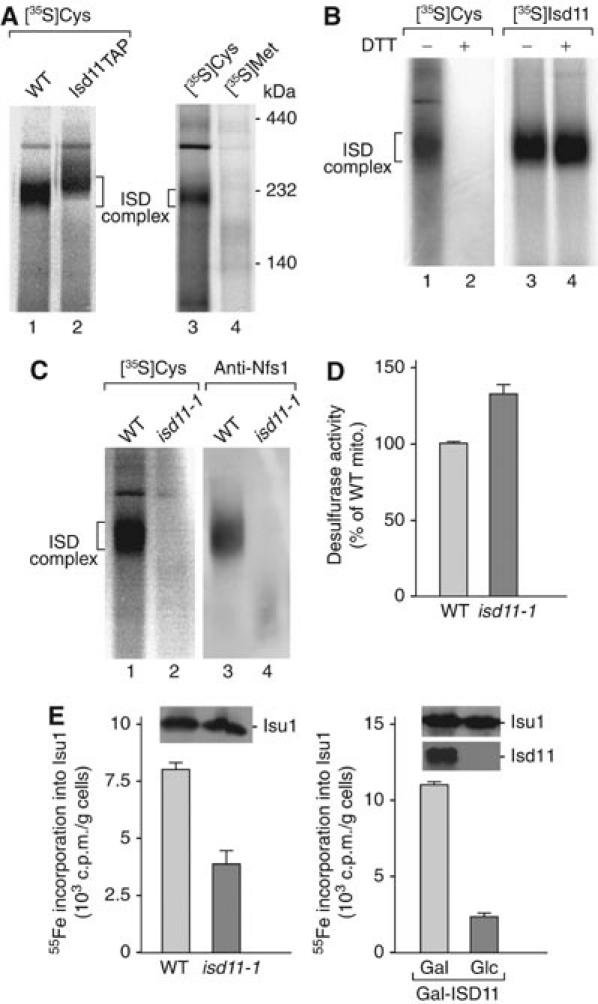
References
-
- Chacinska A, Lind M, Frazier AE, Dudek J, Meisinger C, Geissler A, Sickmann A, Meyer HE, Truscott KN, Guiard B, Pfanner N, Rehling P (2005) Mitochondrial presequence translocase: switching between TOM tethering and motor recruitment involves Tim21 and Tim17. Cell 120: 817–829 - PubMed
-
- Chen OS, Crisp RJ, Valachovic M, Bard M, Winge DR, Kaplan J (2004) Transcription of the yeast iron regulon does not respond directly to iron but rather to iron–sulfur cluster biosynthesis. J Biol Chem 279: 29513–29518 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
