

Stimulation of cannabinoid receptor 2 (CB2) suppresses microglial activation
- PMID: 16343349
- PMCID: PMC1352348
- DOI: 10.1186/1742-2094-2-29
Stimulation of cannabinoid receptor 2 (CB2) suppresses microglial activation
Abstract
Background: Activated microglial cells have been implicated in a number of neurodegenerative disorders, including Alzheimer's disease (AD), multiple sclerosis (MS), and HIV dementia. It is well known that inflammatory mediators such as nitric oxide (NO), cytokines, and chemokines play an important role in microglial cell-associated neuron cell damage. Our previous studies have shown that CD40 signaling is involved in pathological activation of microglial cells. Many data reveal that cannabinoids mediate suppression of inflammation in vitro and in vivo through stimulation of cannabinoid receptor 2 (CB2).
Methods: In this study, we investigated the effects of a cannabinoid agonist on CD40 expression and function by cultured microglial cells activated by IFN-gamma using RT-PCR, Western immunoblotting, flow cytometry, and anti-CB2 small interfering RNA (siRNA) analyses. Furthermore, we examined if the stimulation of CB2 could modulate the capacity of microglial cells to phagocytise Abeta1-42 peptide using a phagocytosis assay.
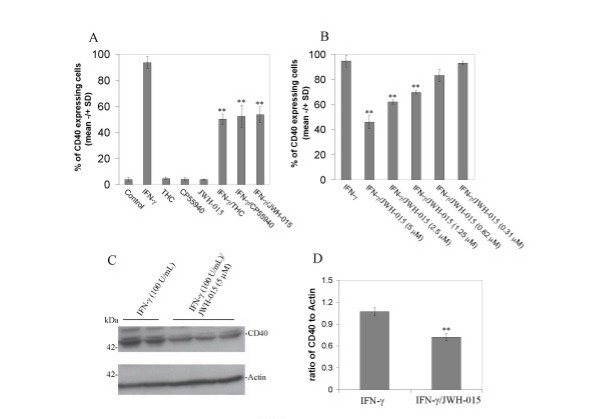
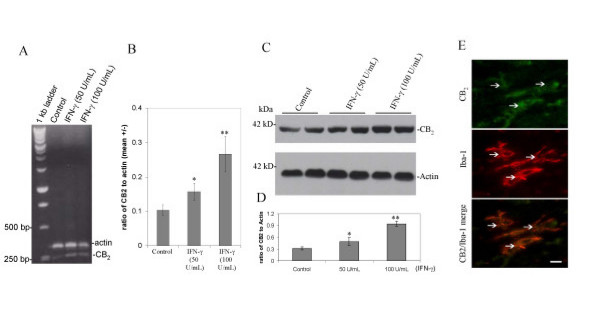
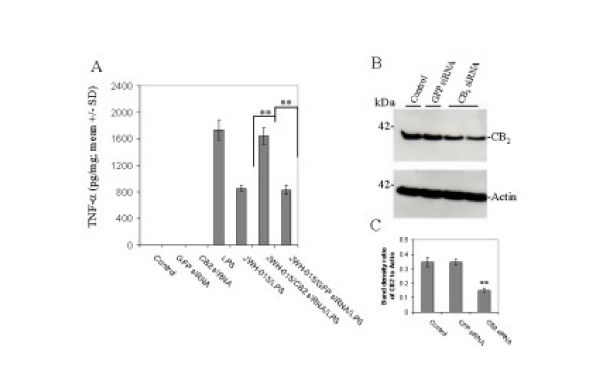
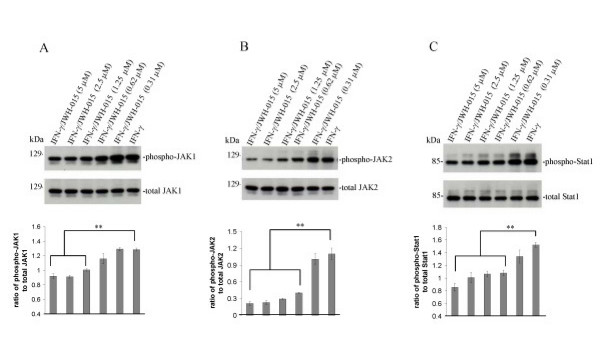
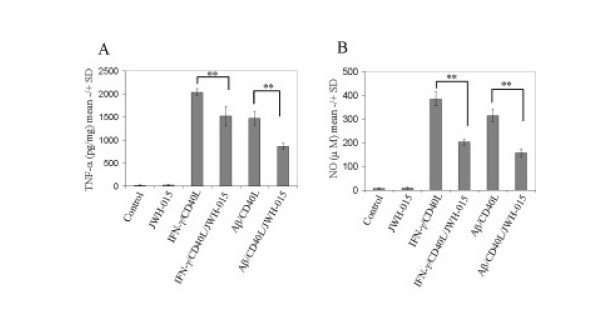
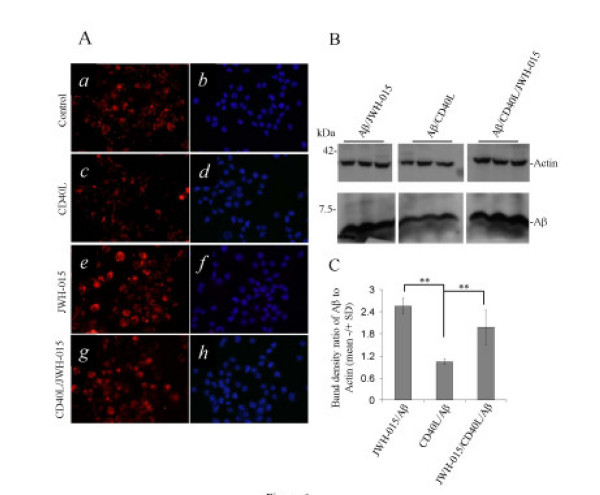
Results: We found that the selective stimulation of cannabinoid receptor CB2 by JWH-015 suppressed IFN-gamma-induced CD40 expression. In addition, this CB2 agonist markedly inhibited IFN-gamma-induced phosphorylation of JAK/STAT1. Further, this stimulation was also able to suppress microglial TNF-alpha and nitric oxide production induced either by IFN-gamma or Abeta peptide challenge in the presence of CD40 ligation. Finally, we showed that CB2 activation by JWH-015 markedly attenuated CD40-mediated inhibition of microglial phagocytosis of Abeta1-42 peptide. Taken together, these results provide mechanistic insight into beneficial effects provided by cannabinoid receptor CB2 modulation in neurodegenerative diseases, particularly AD.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
