

The beta-glucan receptor dectin-1 recognizes specific morphologies of Aspergillus fumigatus
- PMID: 16344862
- PMCID: PMC1311140
- DOI: 10.1371/journal.ppat.0010042
The beta-glucan receptor dectin-1 recognizes specific morphologies of Aspergillus fumigatus
Abstract
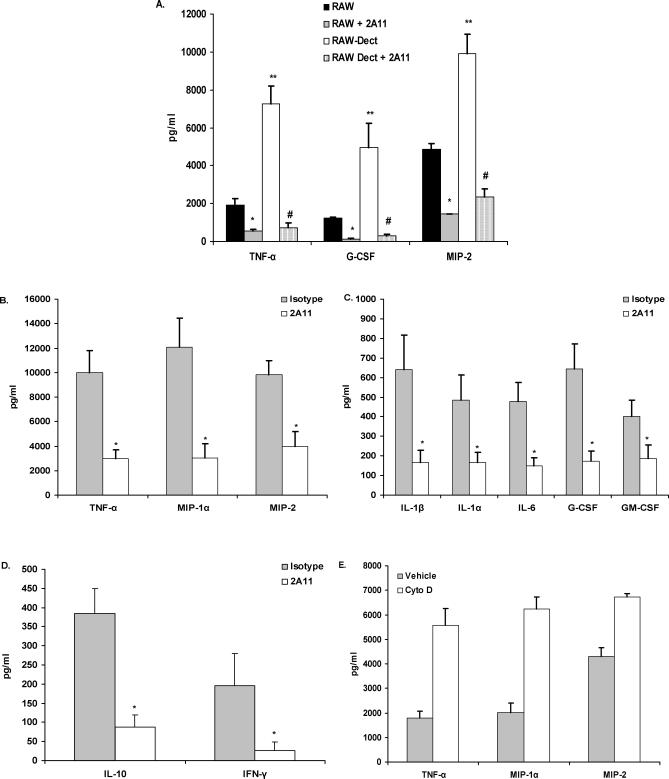
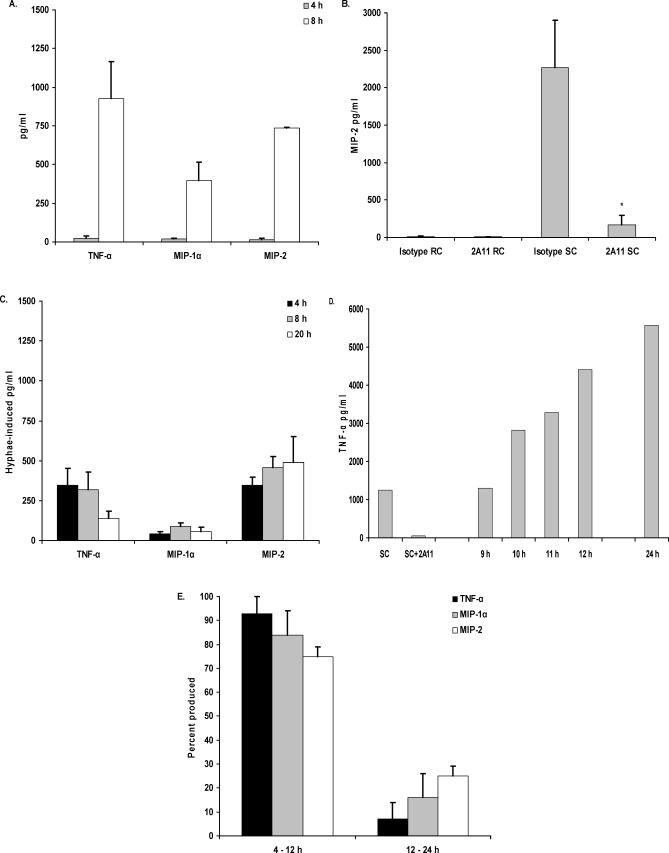
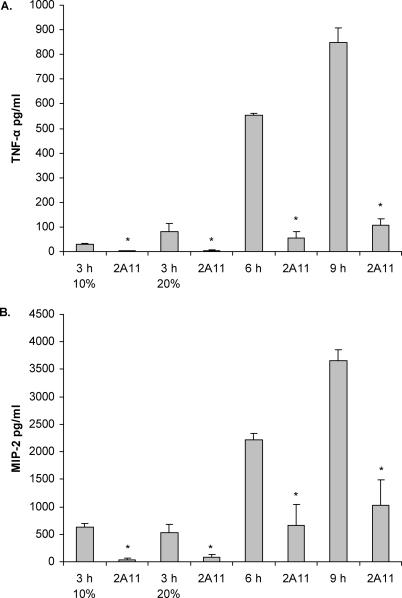
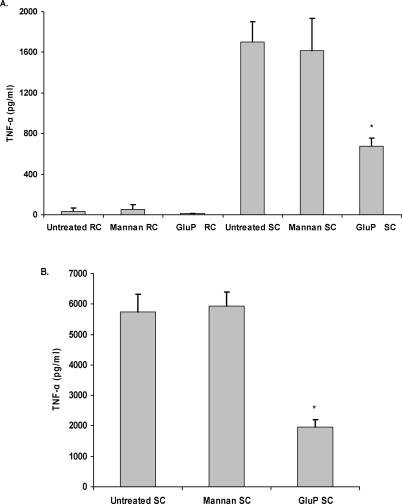
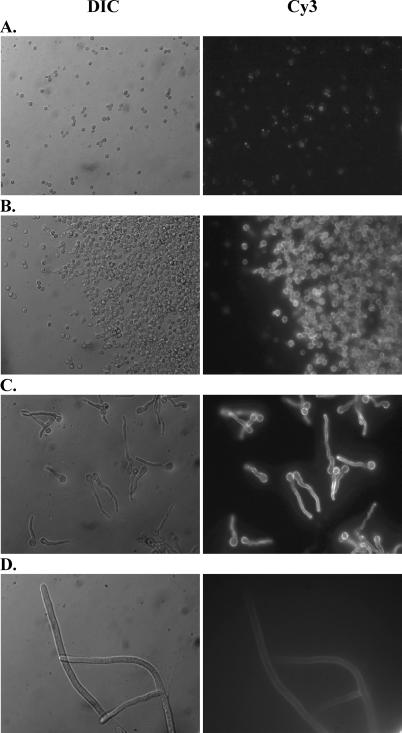
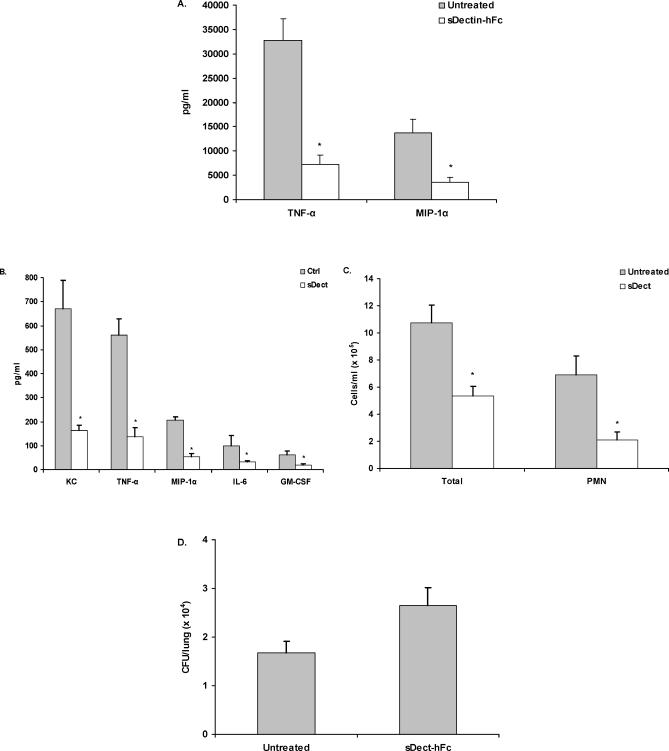
Alveolar macrophages represent a first-line innate host defense mechanism for clearing inhaled Aspergillus fumigatus from the lungs, yet contradictory data exist as to which alveolar macrophage recognition receptor is critical for innate immunity to A. fumigatus. Acknowledging that the A. fumigatus cell wall contains a high beta-1,3-glucan content, we questioned whether the beta-glucan receptor dectin-1 played a role in this recognition process. Monoclonal antibody, soluble receptor, and competitive carbohydrate blockage indicated that the alveolar macrophage inflammatory response, specifically the production of tumor necrosis factor-alpha (TNF-alpha), interleukin-1alpha (IL-1alpha), IL-1beta, IL-6, CXCL2/macrophage inflammatory protein-2 (MIP-2), CCL3/macrophage inflammatory protein-1alpha (MIP-1alpha), granulocyte-colony stimulating factor (G-CSF), and granulocyte monocyte-CSF (GM-CSF), to live A. fumigatus was dependent on recognition via the beta-glucan receptor dectin-1. The inflammatory response was triggered at the highest level by A. fumigatus swollen conidia and early germlings and correlated to the levels of surface-exposed beta glucans, indicating that dectin-1 preferentially recognizes specific morphological forms of A. fumigatus. Intratracheal administration of A. fumigatus conidia to mice in the presence of a soluble dectin-Fc fusion protein reduced both lung proinflammatory cytokine/chemokine levels and cellular recruitment while modestly increasing the A. fumigatus fungal burden, illustrating the importance of beta-glucan-initiated dectin-1 signaling in defense against this pathogen. Collectively, these data show that dectin-1 is centrally required for the generation of alveolar macrophage proinflammatory responses to A. fumigatus and to our knowledge provides the first in vivo evidence for the role of dectin-1 in fungal innate defense.
Conflict of interest statement
Figures
References
-
- Kontoyiannis DP, Bodey GP. Invasive aspergillosis in 2002: An update. Eur J Clin Microbiol Infect Dis. 2002;21:161–172. - PubMed
-
- Baddley JW, Stroud TP, Salzman D, Pappas PG. Invasive mold infections in allogeneic bone marrow transplant recipients. Clin Infect Dis. 2001;32:1319–1324. - PubMed
-
- Denning DW. Invasive aspergillosis. Clin Infect Dis. 1998;26:781–803. - PubMed
-
- Patterson TF, Kirkpatrick WR, White M, Hiemenz JW, Wingard JR, et al. Invasive aspergillosis. Disease spectrum, treatment practices, and outcomes. I3 Aspergillus Study Group. Medicine. 2000;79:250–260. - PubMed
-
- Marr KA, Patterson T, Denning D. Aspergillosis pathogenesis, clinical manifestations and therapy. Infect Dis Clin North Am. 2002;16:875–894. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
