

Herpes simplex virus DNA synthesis is not a decisive regulatory event in the initiation of lytic viral protein expression in neurons in vivo during primary infection or reactivation from latency
- PMID: 16352529
- PMCID: PMC1317510
- DOI: 10.1128/JVI.80.1.38-50.2006
Herpes simplex virus DNA synthesis is not a decisive regulatory event in the initiation of lytic viral protein expression in neurons in vivo during primary infection or reactivation from latency
Abstract
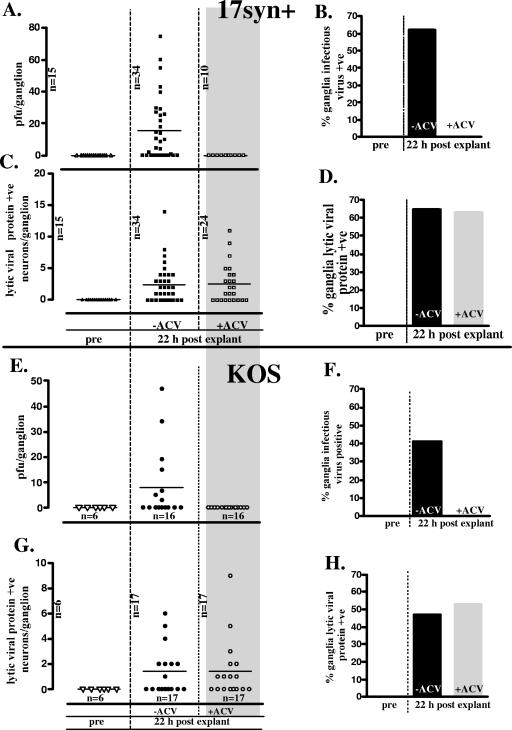
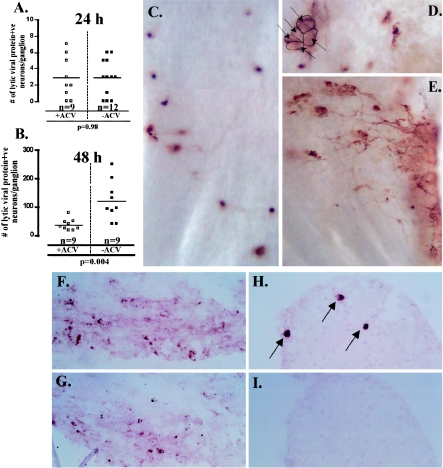
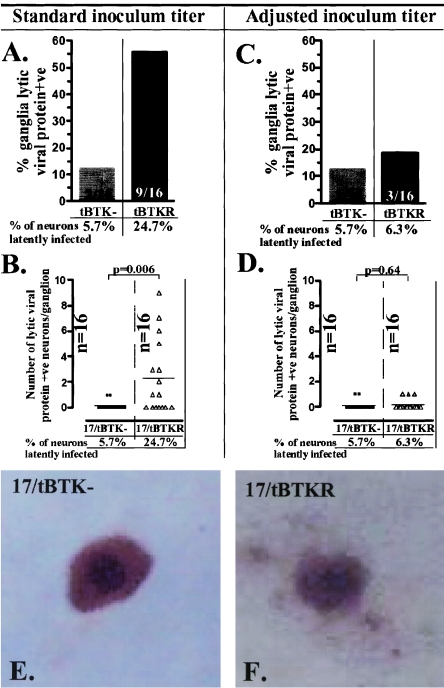
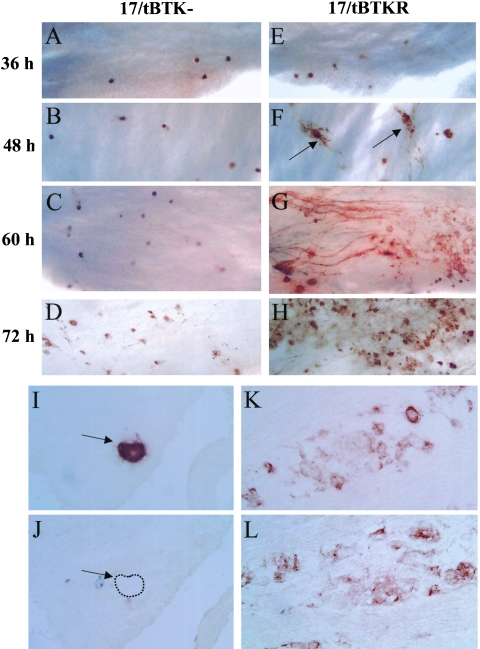
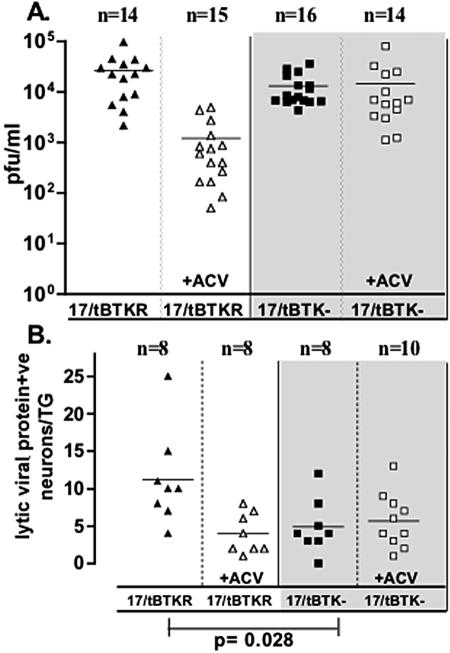
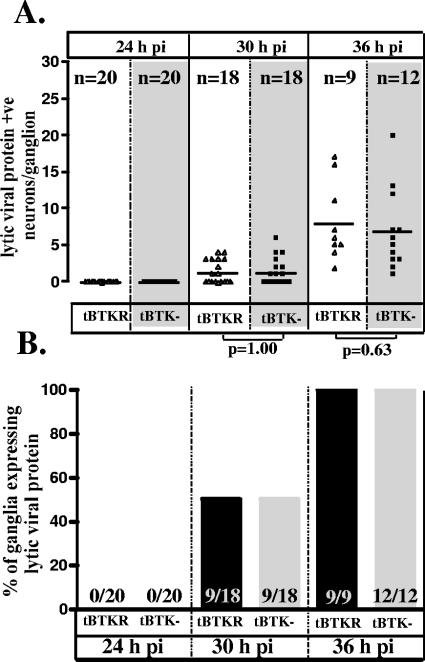
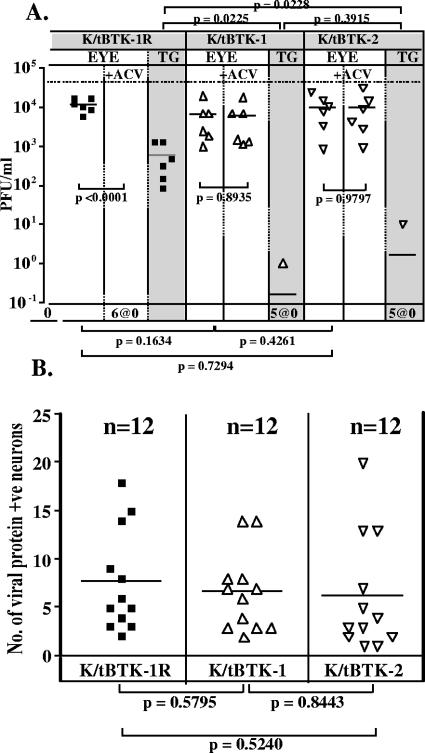
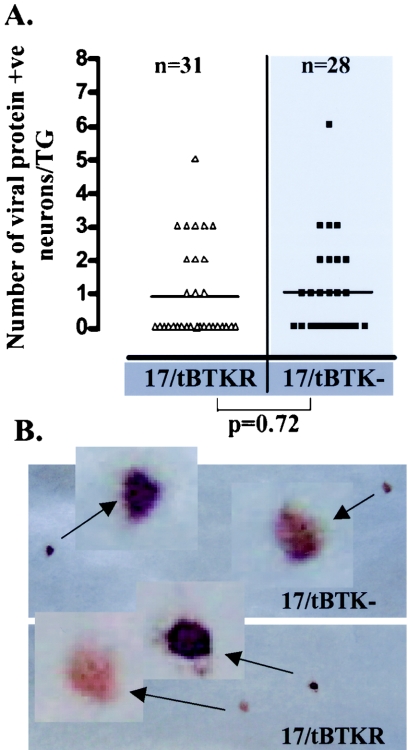
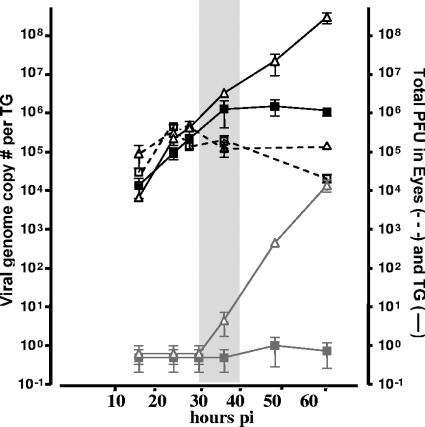
The herpes simplex virus genome can enter a repressed transcriptional state (latency) in sensory neurons of the host nervous system. Although reduced permissiveness of the neuronal environment is widely accepted as a causal factor, the molecular pathway(s) directing and maintaining the viral genome in the latent state remains undefined. Over the past decade, the field has been strongly influenced by the observations of Kosz-Vnenchak et al., which have been interpreted to indicate that, in sensory neurons in vivo, a critical level of viral DNA synthesis within the neuron is required for sufficient viral immediate-early (IE) and early (E) gene expression (M. Kosz-Vnenchak, J. Jacobson, D. M. Coen, and D. M. Knipe, J. Virol. 67:5383-5393, 1993). The levels of IE and E genes are, in turn, thought to regulate the decision to enter the lytic cycle or latency. We have reexamined this issue using new strategies for in situ detection and quantification of viral gene expression in whole tissues. Our results using thymidine kinase-null and rescued mutants as well as wild-type strains in conjunction with viral DNA synthesis blockers demonstrate that (i) despite inhibition of viral DNA replication, many neurons express lytic viral proteins, including IE proteins, during acute infection in the ganglion; (ii) at early times postinoculation, the number of neurons expressing viral proteins in the ganglion is not reduced by inhibition of viral DNA replication; and (iii) following a reactivation stimulus, the numbers of neurons and apparent levels of lytic viral proteins, including IE proteins, are not reduced by inhibition of viral DNA replication. We conclude that viral DNA replication in the neuron per se does not regulate IE gene expression or entry into the lytic cycle.
Figures
References
-
- Coen, D. M., M. Kosz-Vnenchak, J. G. Jacobson, D. A. Leib, C. L. Bogard, P. A. Schaffer, K. L. Tyler, and D. M. Knipe. 1989. Thymidine kinase-negative herpes simplex virus mutants establish latency in mouse trigeminal ganglia but do not reactivate. Proc. Natl. Acad. Sci. USA 86:4736-4740. - PMC - PubMed
-
- Davido, D. J., and D. A. Leib. 1996. Role of cis-acting sequences of the ICPO promoter of herpes simplex virus type 1 in viral pathogenesis, latency and reactivation. J. Gen. Virol. 77:1853-1863. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
