

Secretory leucoprotease inhibitor binds to NF-kappaB binding sites in monocytes and inhibits p65 binding
- PMID: 16352738
- PMCID: PMC2212970
- DOI: 10.1084/jem.20050768
Secretory leucoprotease inhibitor binds to NF-kappaB binding sites in monocytes and inhibits p65 binding
Abstract
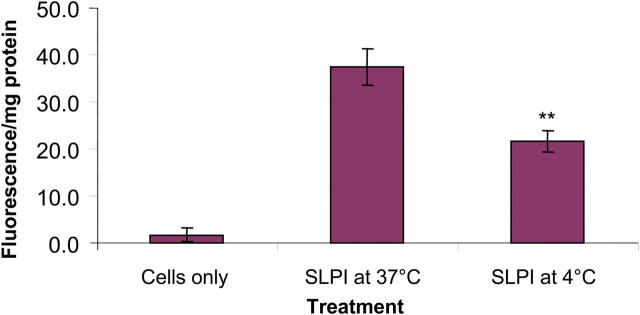
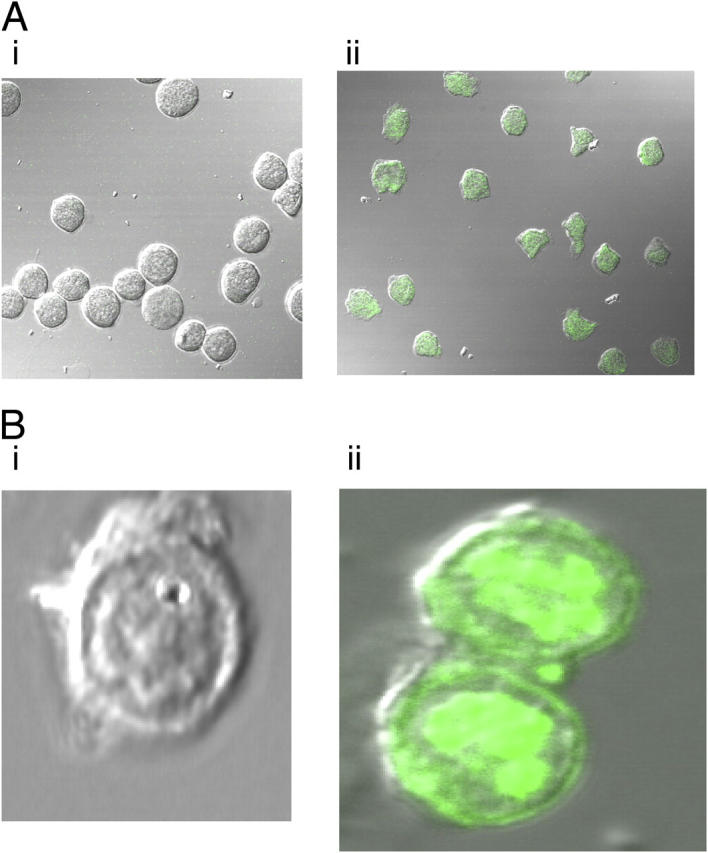
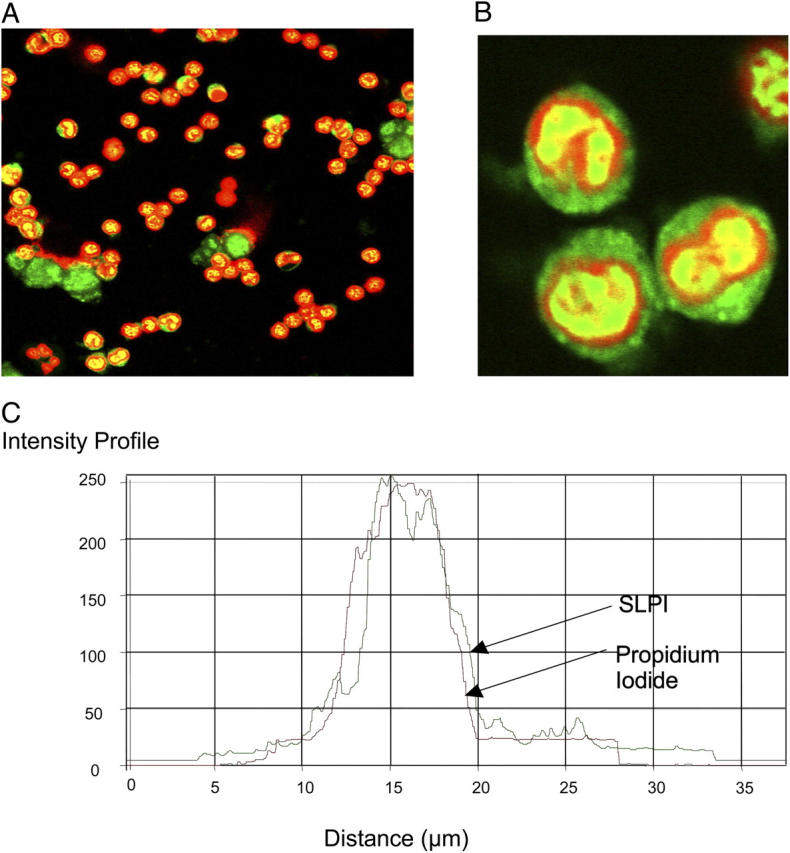
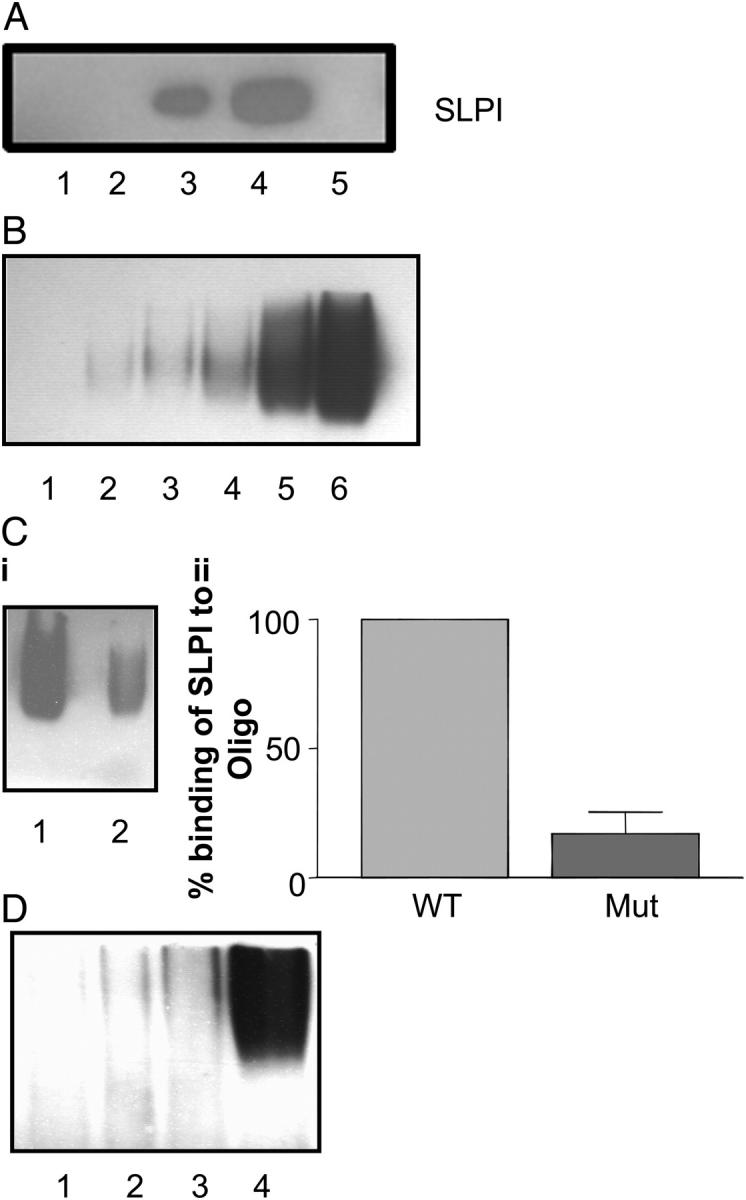
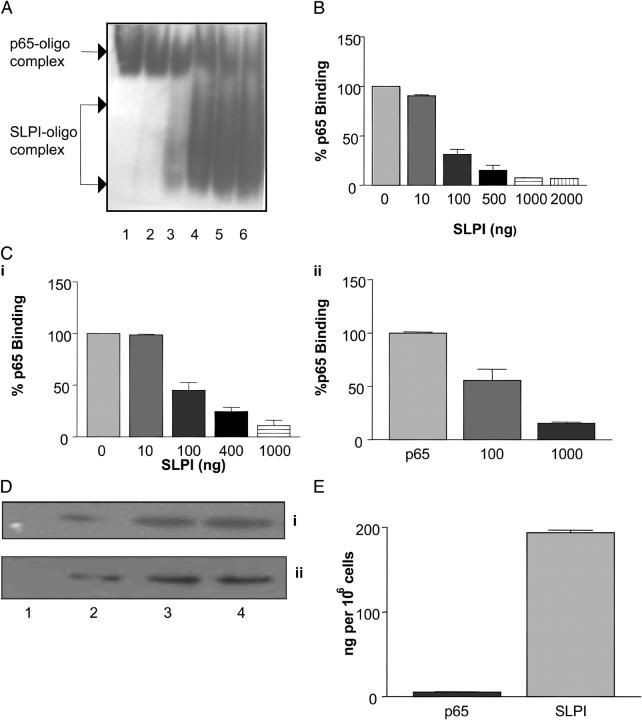
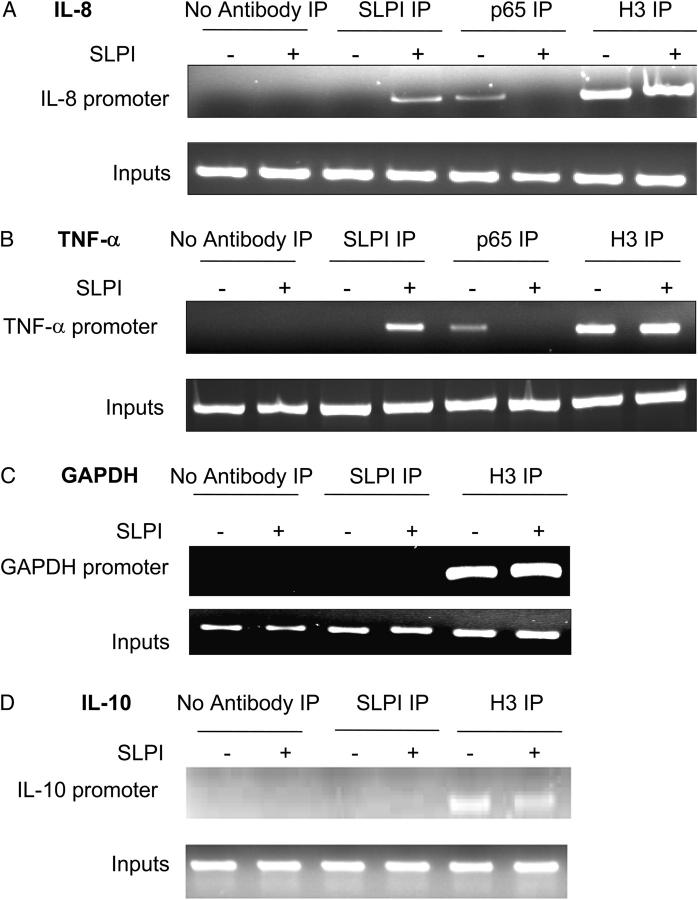
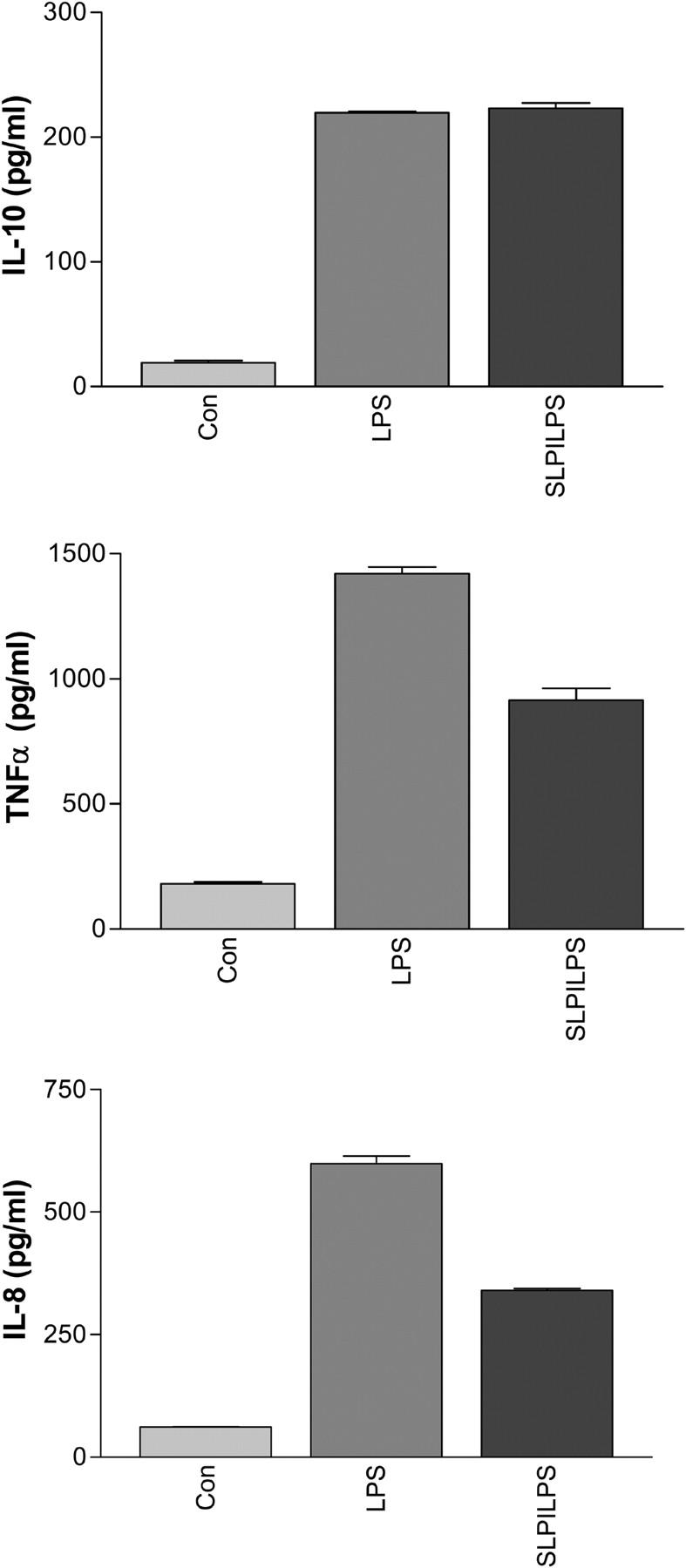
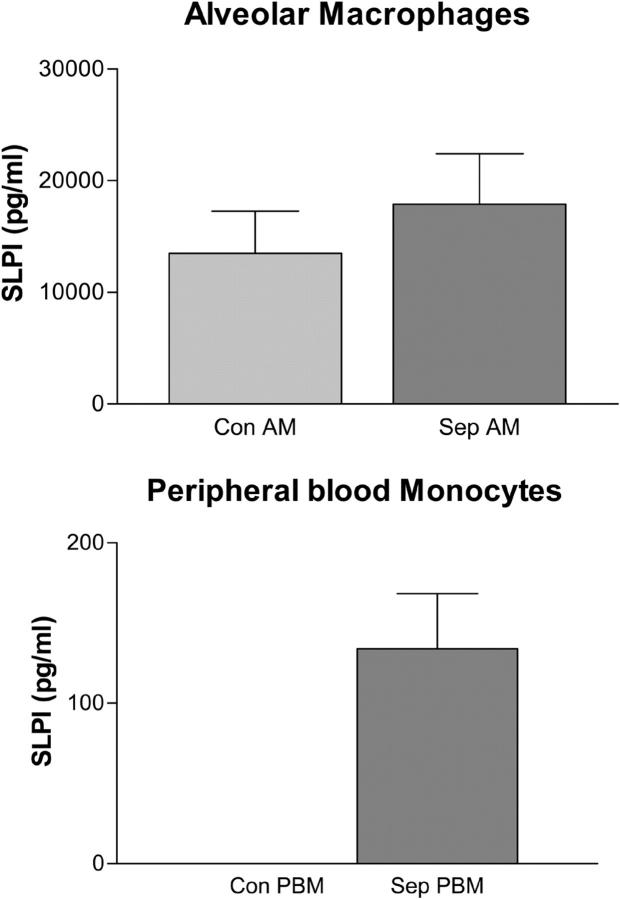
Secretory leucoprotease inhibitor (SLPI) is a nonglycosylated protein produced by epithelial cells. In addition to its antiprotease activity, SLPI has been shown to exhibit antiinflammatory properties, including down-regulation of tumor necrosis factor alpha expression by lipopolysaccharide (LPS) in macrophages and inhibition of nuclear factor (NF)-kappaB activation in a rat model of acute lung injury. We have previously shown that SLPI can inhibit LPS-induced NF-kappaB activation in monocytic cells by inhibiting degradation of IkappaBalpha without affecting the LPS-induced phosphorylation and ubiquitination of IkappaBalpha. Here, we present evidence to show that upon incubation with peripheral blood monocytes (PBMs) and the U937 monocytic cell line, SLPI enters the cells, becoming rapidly localized to the cytoplasm and nucleus, and affects NF-kappaB activation by binding directly to NF-kappaB binding sites in a site-specific manner. SLPI can also prevent p65 interaction with the NF-kappaB consensus region at concentrations commensurate with the physiological nuclear levels of SLPI and p65. We also demonstrate the presence of SLPI in nuclear fractions of PBMs and alveolar macrophages from individuals with cystic fibrosis and community-acquired pneumonia. Therefore, SLPI inhibition of NF-kappaB activation is mediated, in part, by competitive binding to the NF-kappaB consensus-binding site.
Figures

References
-
- Gauthier, F., U. Fryksmark, K. Ohlsson, and J.G. Bieth. 1982. Kinetics of the inhibition of leukocyte elastase by the bronchial inhibitor. Biochim. Biophys. Acta. 700:178–183. - PubMed
-
- Seemuller, U., M. Arnhold, H. Fritz, K. Wiedenmann, W. Machleidt, R. Heinzel, H. Applehans, H.G. Gassen, and F. Lottspeich. 1986. The acid-stable proteinase inhibitor of human mucous secretions (HUSI-I, antileukoprotease). Complete amino acid sequence as revealed by protein and cDNA sequencing and structural homology to whey proteins and Red Sea turtle proteinase inhibitor. FEBS Lett. 199:43–48. - PubMed
-
- Hiemstra, P.S., B.A. Fernie-King, J. McMichael, P.J. Lachmann, and J.M. Sallenave. 2004. Antimicrobial peptides: mediators of innate immunity as templates for the development of novel anti-infective and immune therapeutics. Curr. Pharm. Des. 10:2891–2905. - PubMed
-
- Jin, F.Y., C. Nathan, D. Radzioch, and A. Ding. 1997. Secretory leukocyte protease inhibitor: a macrophage product induced by and antagonistic to bacterial lipopolysaccharide. Cell. 88:417–426. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
