

A C-terminal domain directs Kv3.3 channels to dendrites
- PMID: 16354911
- PMCID: PMC6726014
- DOI: 10.1523/JNEUROSCI.3672-05.2005
A C-terminal domain directs Kv3.3 channels to dendrites
Abstract
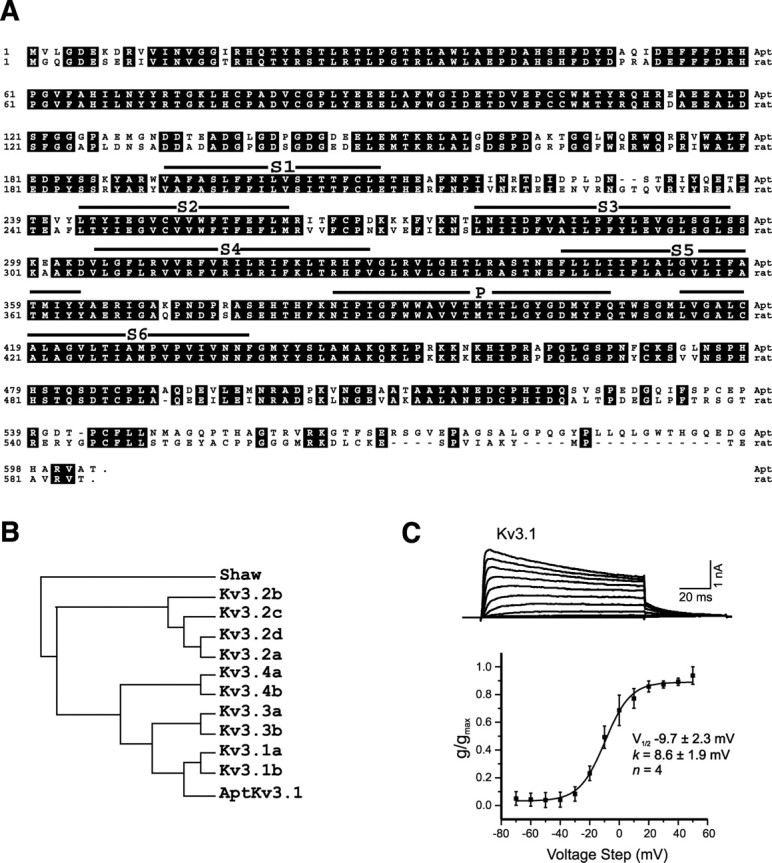
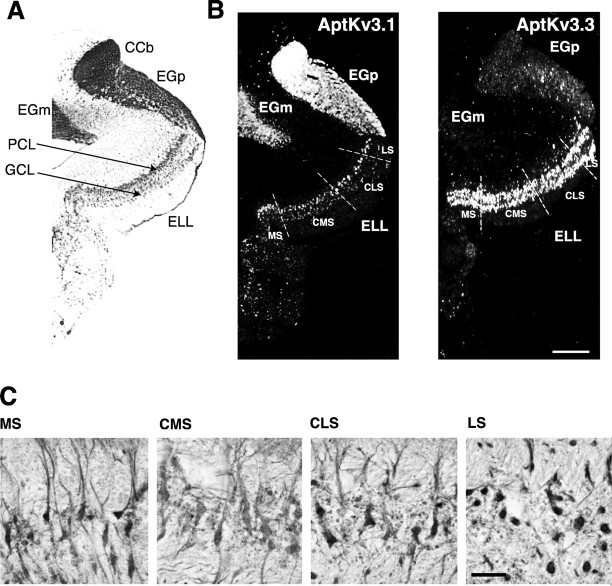
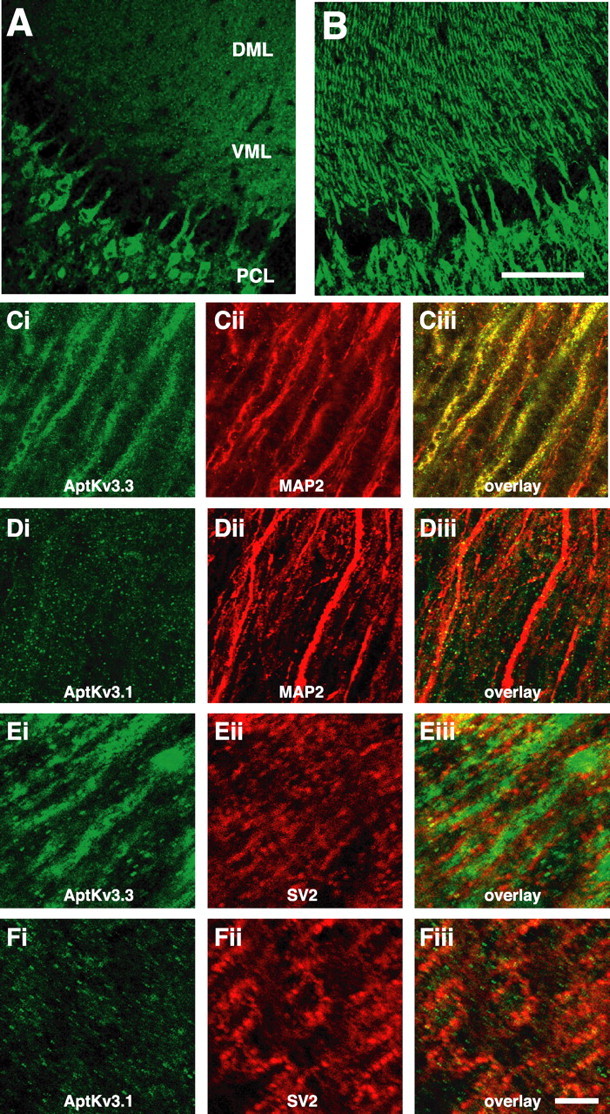
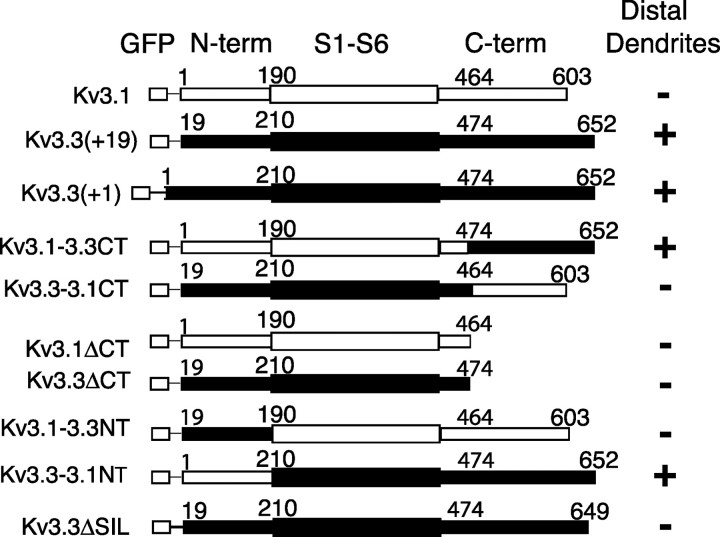
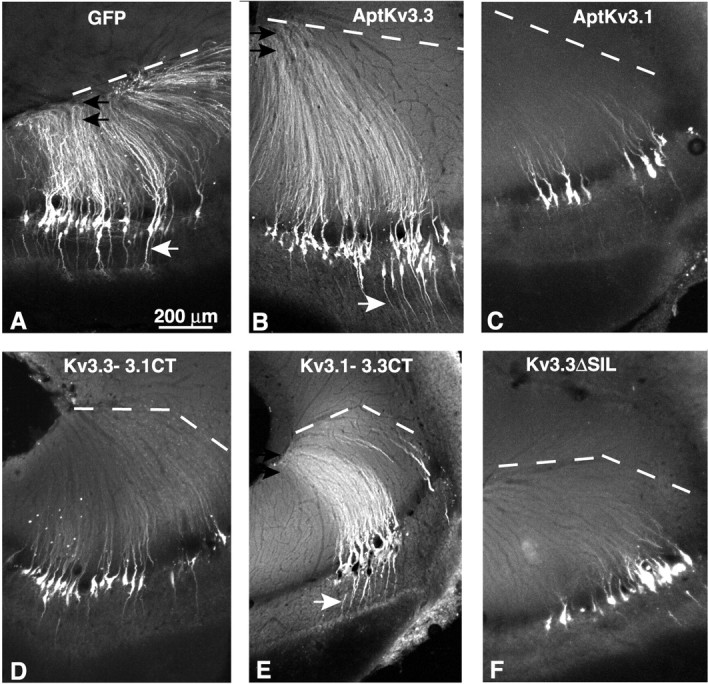
Pyramidal neurons of the electrosensory lateral line lobe (ELL) of Apteronotus leptorhynchus express Kv3-type voltage-gated potassium channels that give rise to high-threshold currents at the somatic and dendritic levels. Two members of the Kv3 channel family, AptKv3.1 and AptKv3.3, are coexpressed in these neurons. AptKv3.3 channels are expressed at uniformly high levels in each of four ELL segments, whereas AptKv3.1 channels appear to be expressed in a graded manner with higher levels of expression in segments that process high-frequency electrosensory signals. Immunohistochemical and recombinant channel expression studies show a differential distribution of these two channels in the dendrites of ELL pyramidal neurons. AptKv3.1 is concentrated in somas and proximal dendrites, whereas AptKv3.3 is distributed throughout the full extent of the large dendritic tree. Recombinant channel expression of AptKv3 channels through in vivo viral injections allowed directed retargeting of AptKv3 subtypes over the somadendritic axis, revealing that the sequence responsible for targeting channels to distal dendrites lies within the C-terminal domain of the AptKv3.3 protein. The targeting domain includes a consensus sequence predicted to bind to a PDZ (postsynaptic density-95/Discs large/zona occludens-1)-type protein-protein interaction motif. These findings reveal that different functional roles for Kv3 potassium channels at the somatic and dendritic level of a sensory neuron are attained through specific targeting that selectively distributes Kv3.3 channels to the dendritic compartment.
Figures
Similar articles
-
The contribution of dendritic Kv3 K+ channels to burst threshold in a sensory neuron.J Neurosci. 2001 Jan 1;21(1):125-35. doi: 10.1523/JNEUROSCI.21-01-00125.2001. J Neurosci. 2001. PMID: 11150328 Free PMC article.
-
A prominent soma-dendritic distribution of Kv3.3 K+ channels in electrosensory and cerebellar neurons.J Comp Neurol. 2001 Dec 17;441(3):234-47. doi: 10.1002/cne.1409. J Comp Neurol. 2001. PMID: 11745647
-
Inactivation of Kv3.3 potassium channels in heterologous expression systems.J Biol Chem. 2003 Oct 17;278(42):40890-8. doi: 10.1074/jbc.M304235200. Epub 2003 Aug 15. J Biol Chem. 2003. PMID: 12923191
-
Oscillatory burst discharge generated through conditional backpropagation of dendritic spikes.J Physiol Paris. 2002 Sep-Dec;96(5-6):517-30. doi: 10.1016/S0928-4257(03)00007-X. J Physiol Paris. 2002. PMID: 14692499 Review.
-
Distribution and function of potassium channels in the electrosensory lateral line lobe of weakly electric apteronotid fish.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2006 Jun;192(6):637-48. doi: 10.1007/s00359-006-0103-z. Epub 2006 Jan 20. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2006. PMID: 16425062 Review.
Cited by
-
The axon-dendrite targeting of Kv3 (Shaw) channels is determined by a targeting motif that associates with the T1 domain and ankyrin G.J Neurosci. 2007 Dec 19;27(51):14158-70. doi: 10.1523/JNEUROSCI.3675-07.2007. J Neurosci. 2007. PMID: 18094255 Free PMC article.
-
Differences in Sodium Channel Densities in the Apical Dendrites of Pyramidal Cells of the Electrosensory Lateral Line Lobe.Front Neural Circuits. 2019 Jun 4;13:41. doi: 10.3389/fncir.2019.00041. eCollection 2019. Front Neural Circuits. 2019. PMID: 31213991 Free PMC article.
-
Polarized targeting of ion channels in neurons.Pflugers Arch. 2007 Mar;453(6):763-9. doi: 10.1007/s00424-006-0155-5. Epub 2006 Nov 8. Pflugers Arch. 2007. PMID: 17091311 Review.
-
Balanced ionotropic receptor dynamics support signal estimation via voltage-dependent membrane noise.J Neurophysiol. 2016 Jan 1;115(1):530-45. doi: 10.1152/jn.00786.2015. Epub 2015 Nov 11. J Neurophysiol. 2016. PMID: 26561607 Free PMC article.
-
Function and mechanism of axonal targeting of voltage-sensitive potassium channels.Prog Neurobiol. 2011 Jul;94(2):115-32. doi: 10.1016/j.pneurobio.2011.04.009. Epub 2011 Apr 22. Prog Neurobiol. 2011. PMID: 21530607 Free PMC article. Review.
References
-
- Arnold DB, Clapham DE (1999) Molecular determinants for subcellular localization of PSD-95 with an interacting K+ channel. Neuron 23: 149–157. - PubMed
-
- Berman NJ, Maler L (1999) Neural architecture of the electrosensory lateral line lobe: adaptations for coincidence detection, a sensory searchlight and frequency-dependent adaptive filtering. J Exp Biol 202: 1243–1253. - PubMed
-
- Carr C, Maler L (1986) Electroreception in Gymnotiform fish. In: Electroreception (Bullock TH, Heiligenberg W, eds), pp 319–373. New York: Wiley.
-
- Chenchik A, Diachenko L, Moqadam F, Tarabykin V, Lukyanov S, Siebert PD (1996) Full-length cDNA cloning and determination of mRNA 5′ and 3′ ends by amplification of adaptor-ligated cDNA. Biotechniques 21: 526–534. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources