

BACE1, a major determinant of selective vulnerability of the brain to amyloid-beta amyloidogenesis, is essential for cognitive, emotional, and synaptic functions
- PMID: 16354928
- PMCID: PMC2564291
- DOI: 10.1523/JNEUROSCI.2766-05.2005
BACE1, a major determinant of selective vulnerability of the brain to amyloid-beta amyloidogenesis, is essential for cognitive, emotional, and synaptic functions
Abstract
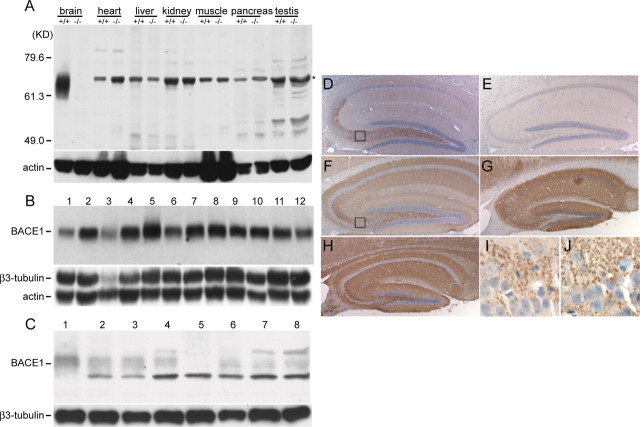
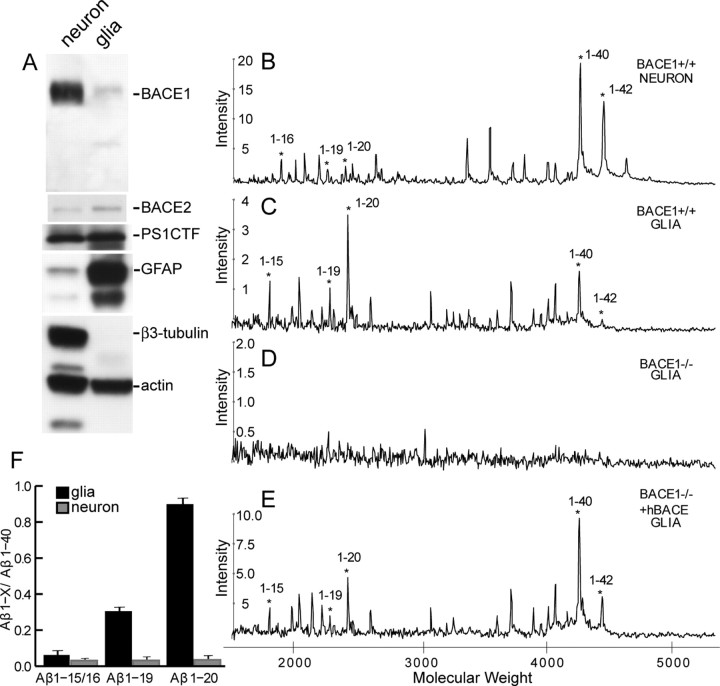
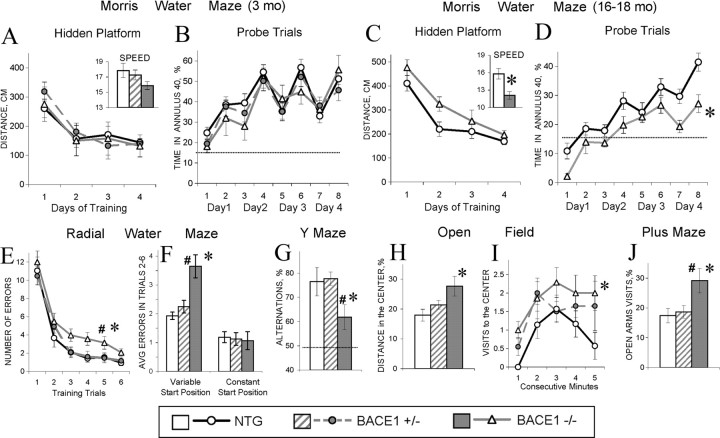
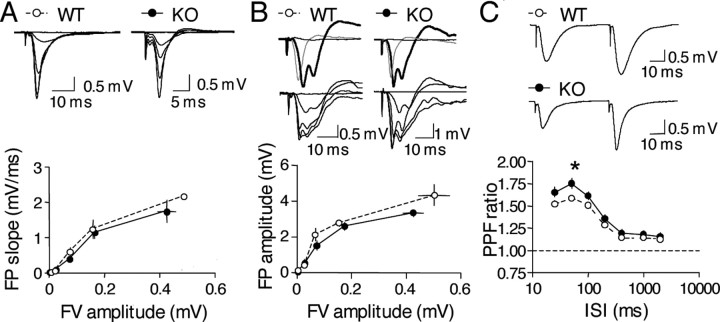
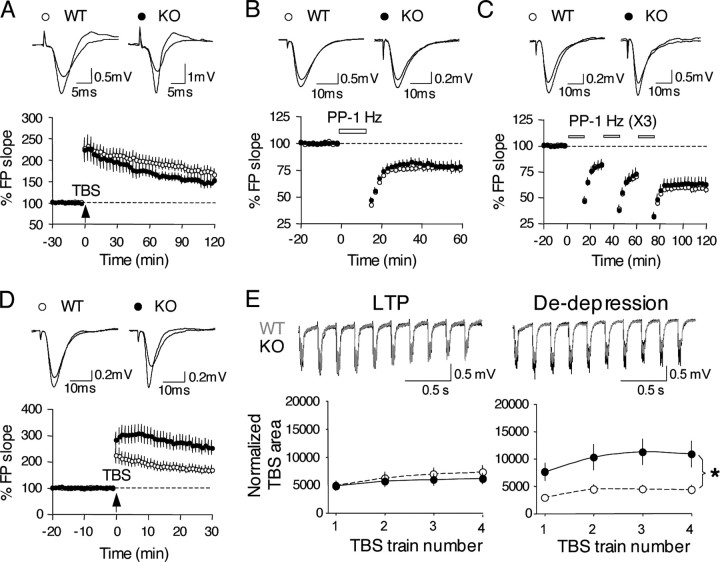
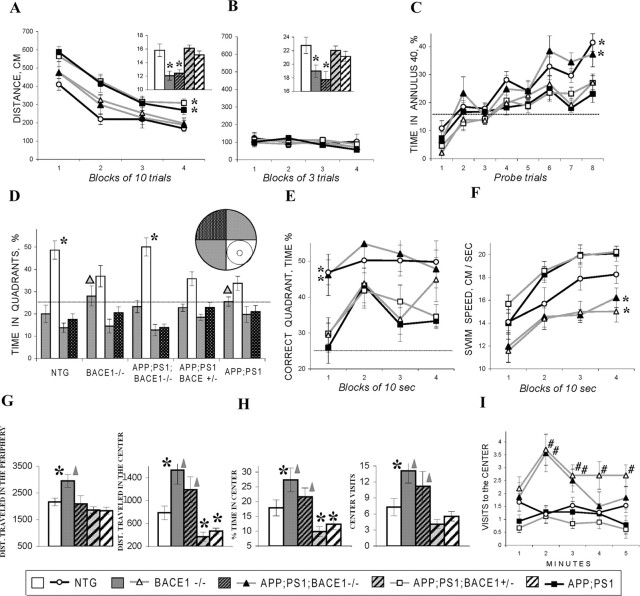
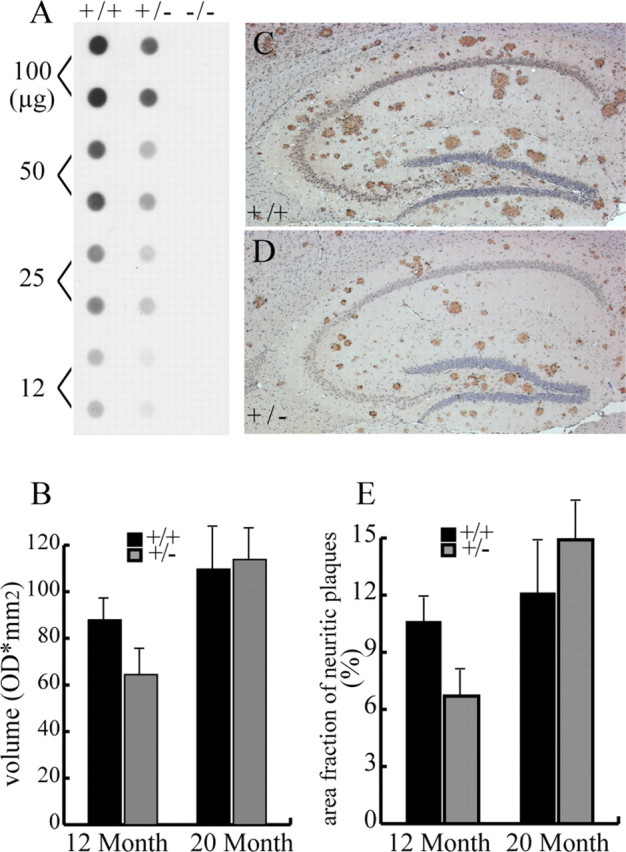
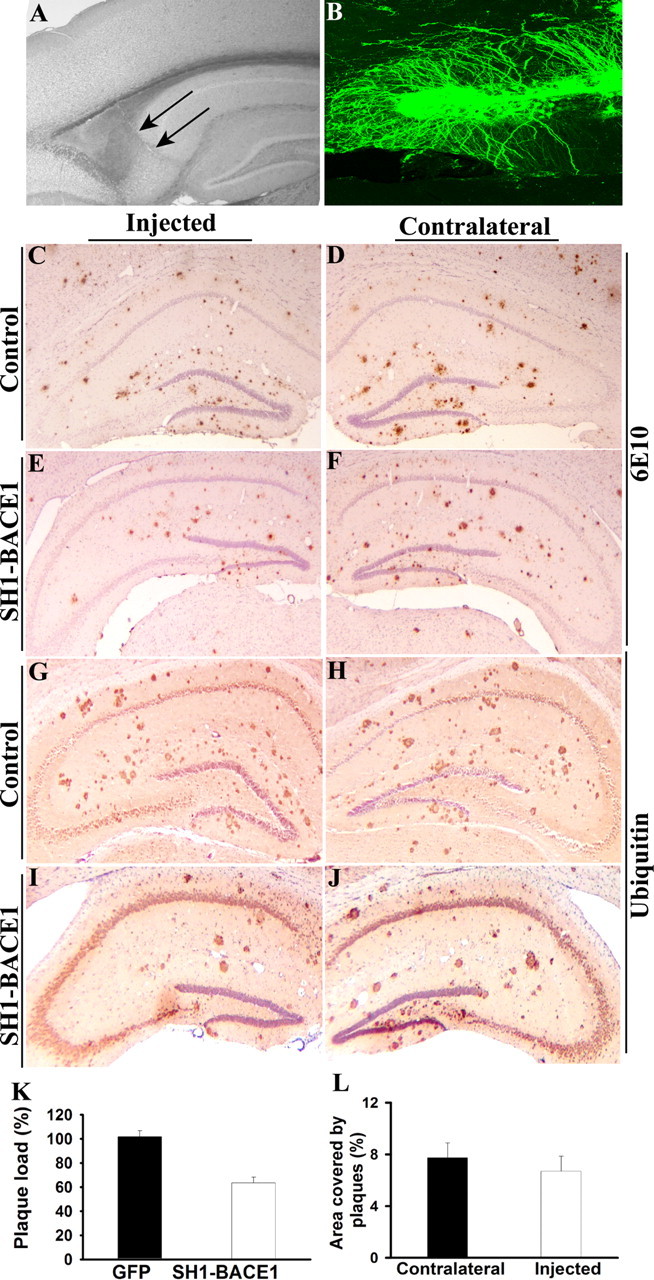
A transmembrane aspartyl protease termed beta-site APP cleavage enzyme 1 (BACE1) that cleaves the amyloid-beta precursor protein (APP), which is abundant in neurons, is required for the generation of amyloid-beta (Abeta) peptides implicated in the pathogenesis of Alzheimer's disease (AD). We now demonstrate that BACE1, enriched in neurons of the CNS, is a major determinant that predisposes the brain to Abeta amyloidogenesis. The physiologically high levels of BACE1 activity coupled with low levels of BACE2 and alpha-secretase anti-amyloidogenic activities in neurons is a major contributor to the accumulation of Abeta in the CNS, whereas other organs are spared. Significantly, deletion of BACE1 in APPswe;PS1DeltaE9 mice prevents both Abeta deposition and age-associated cognitive abnormalities that occur in this model of Abeta amyloidosis. Moreover, Abeta deposits are sensitive to BACE1 dosage and can be efficiently cleared from the CNS when BACE1 is silenced. However, BACE1 null mice manifest alterations in hippocampal synaptic plasticity as well as in performance on tests of cognition and emotion. Importantly, memory deficits but not emotional alterations in BACE1(-/-) mice are prevented by coexpressing APPswe;PS1DeltaE9 transgenes, indicating that other potential substrates of BACE1 may affect neural circuits related to emotion. Our results establish BACE1 and APP processing pathways as critical for cognitive, emotional, and synaptic functions, and future studies should be alert to potential mechanism-based side effects that may occur with BACE1 inhibitors designed to ameliorate Abeta amyloidosis in AD.
Figures
References
-
- Abeliovich A, Chen C, Goda Y, Silva AJ, Stevens CF, Tonegawa S (1993) Modified hippocampal long-term potentiation in PKC gamma-mutant mice. Cell 75: 1253–1262. - PubMed
-
- Allinson TM, Parkin ET, Turner AJ, Hooper NM (2003) ADAMs family members as amyloid precursor protein alpha-secretases. J Neurosci Res 74: 342–352. - PubMed
-
- Arendash GW, King DL, Gordon MN, Morgan D, Hatcher JM, Hope CE, Diamond DM (2001) Progressive, age-related behavioral impairments in transgenic mice carrying both mutant amyloid precursor protein and presenilin-1 transgenes. Brain Res 891: 42–53. - PubMed
-
- Ashe KH (2001) Learning and memory in transgenic mice modeling Alzheimer's disease. Learn Mem 8: 301–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases