

Spinal cord-transected mice learn to step in response to quipazine treatment and robotic training
- PMID: 16354932
- PMCID: PMC6726027
- DOI: 10.1523/JNEUROSCI.1523-05.2005
Spinal cord-transected mice learn to step in response to quipazine treatment and robotic training
Abstract
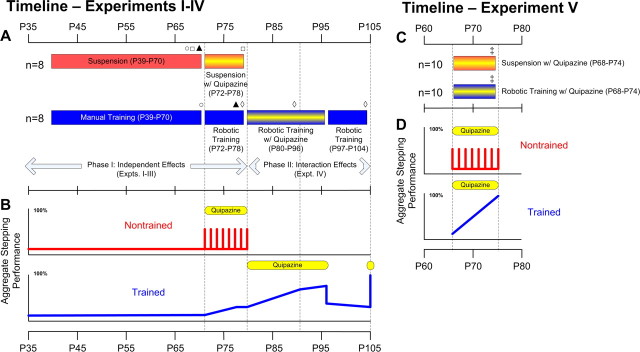
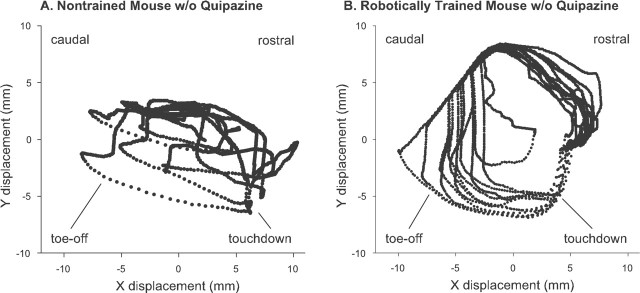
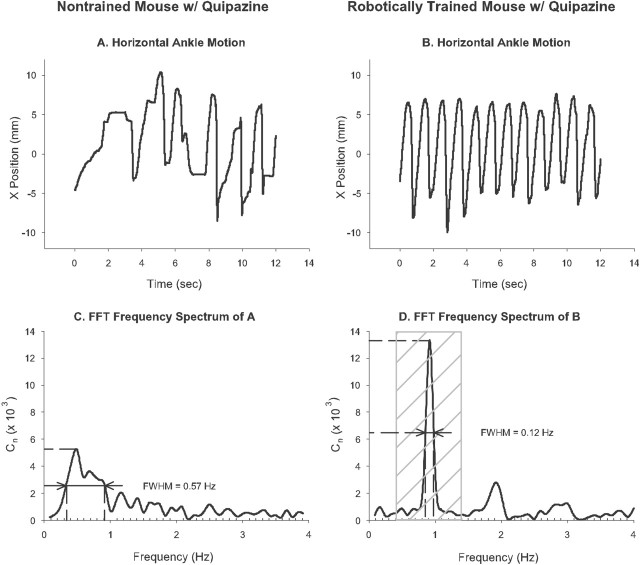
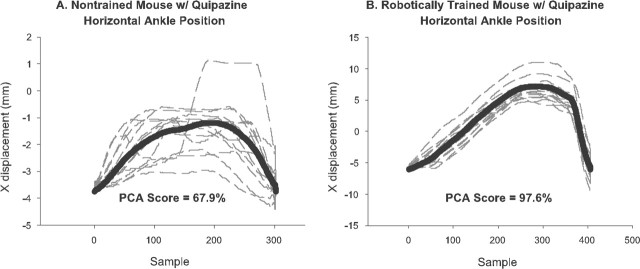
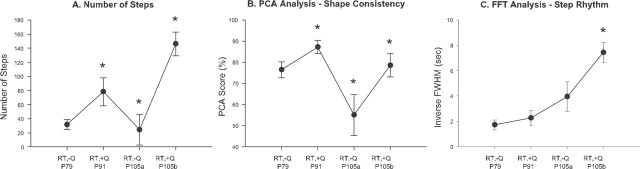
In the present study, concurrent treatment with robotic step training and a serotonin agonist, quipazine, generated significant recovery of locomotor function in complete spinal cord-transected mice (T7-T9) that otherwise could not step. The extent of recovery achieved when these treatments were combined exceeded that obtained when either treatment was applied independently. We quantitatively analyzed the stepping characteristics of spinal mice after alternatively administering no training, manual training, robotic training, quipazine treatment, or a combination of robotic training with quipazine treatment, to examine the mechanisms by which training and quipazine treatment promote functional recovery. Using fast Fourier transform and principal components analysis, significant improvements in the step rhythm, step shape consistency, and number of weight-bearing steps were observed in robotically trained compared with manually trained or nontrained mice. In contrast, manual training had no effect on stepping performance, yielding no improvement compared with nontrained mice. Daily bolus quipazine treatment acutely improved the step shape consistency and number of steps executed by both robotically trained and nontrained mice, but these improvements did not persist after quipazine was withdrawn. At the dosage used (0.5 mg/kg body weight), quipazine appeared to facilitate, rather than directly generate, stepping, by enabling the spinal cord neural circuitry to process specific patterns of sensory information associated with weight-bearing stepping. Via this mechanism, quipazine treatment enhanced kinematically appropriate robotic training. When administered intermittently during an extended period of robotic training, quipazine revealed training-induced stepping improvements that were masked in the absence of the pharmacological treatment.
Figures
References
-
- Anden NE, Haggendal J, Magnusson T, Rodengren E (1964) The time course of the disappearance of noradrenaline and 5-hydroxtryptamine in the spinal cord after transection. Acta Physiol 62: 115–118. - PubMed
-
- Antri M, Orsal D, Barthe JY (2002) Locomotor recovery in the chronic spinal rat: effects of long-term treatment with a 5-HT2 agonist. Eur J Neurosci 16: 467–476. - PubMed
-
- Barbeau H, Rossignol S (1990) The effects of serotonergic drugs on the locomotor pattern and on cutaneous reflexes of the adult chronic spinal cat. Brain Res 514: 55–67. - PubMed
-
- Barbeau H, Rossignol S (1991) Initiation and modulation of the locomotor pattern in the adult chronic spinal cat by noradrenergic, serotonergic and dopaminergic drugs. Brain Res 546: 250–260. - PubMed
-
- Behrman AL, Harkema SJ (2000) Locomotor training after human spinal cord injury: a series of case studies. Phys Ther 80: 688–700. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical