

Probing ion-channel pores one proton at a time
- PMID: 16355215
- PMCID: PMC1384014
- DOI: 10.1038/nature04293
Probing ion-channel pores one proton at a time
Abstract
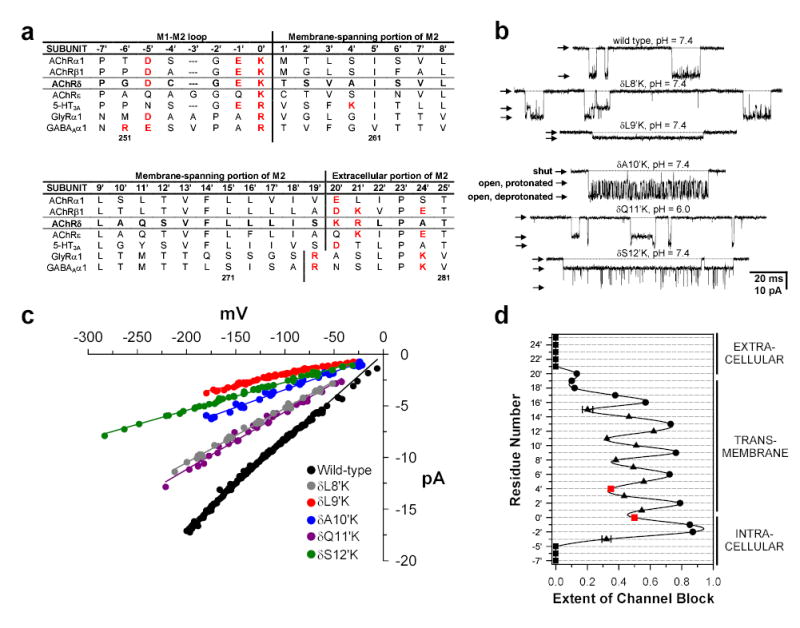
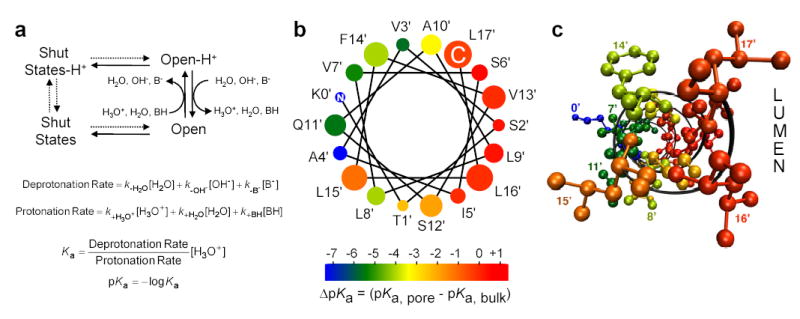
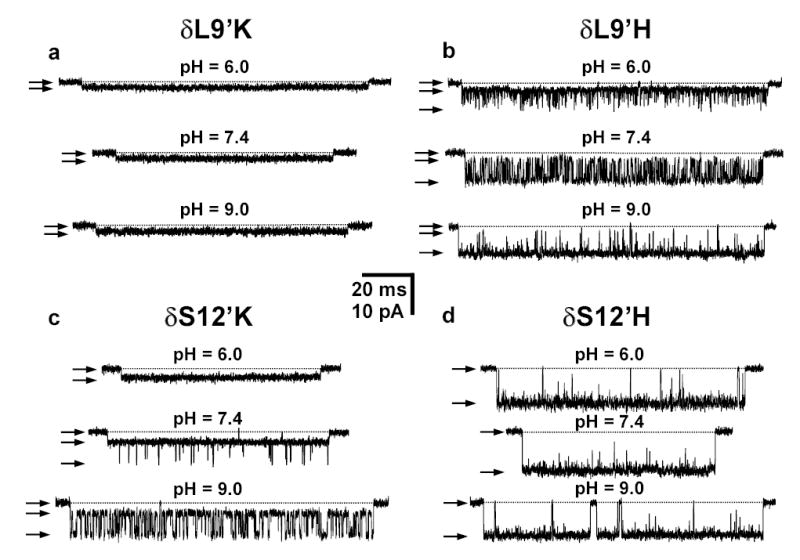
Although membrane proteins often rely on ionizable residues for structure and function, their ionization states under physiological conditions largely elude experimental estimation. To gain insight into the effect of the local microenvironment on the proton affinity of ionizable residues, we have engineered individual lysines, histidines and arginines along the alpha-helical lining of the transmembrane pore of the nicotinic acetylcholine receptor. We can detect individual proton binding-unbinding reactions electrophysiologically at the level of a single proton on a single side chain as brief blocking-unblocking events of the passing cation current. Kinetic analysis of these fluctuations yields the position-dependent rates of proton transfer, from which the corresponding pK(a) values and shifts in pK(a) can be calculated. Here we present a self-consistent, residue-by-residue description of the microenvironment around the pore-lining transmembrane alpha-helices (M2) in the open-channel conformation, in terms of the excess free energy that is required to keep the engineered basic side chains protonated relative to bulk water. A comparison with closed-channel data leads us to propose that the rotation of M2, which is frequently invoked as a hallmark of the gating mechanism of Cys-loop receptors, is minimal, if any.
Figures
References
-
- Perutz MF. Electrostatic effects in proteins. Science. 1978;201:1187–1191. - PubMed
-
- Warshel A. Electrostatic basis of structure-function correlation in proteins. Acc Chem Res. 1981;14:284–290.
-
- Davis ME, McCammon JA. Electrostatics in biomolecular structure and dynamics. Chem Rev. 1990;90:509–521.
-
- Honig B, Nicholls A. Classical electrostatics in biology and chemistry. Science. 1995;268:1144–1149. - PubMed
-
- Imoto K, et al. Rings of negatively charged amino acids determine the acetylcholine receptor channel conductance. Nature. 1988;335:645–648. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
