

Role for hypocretin in mediating stress-induced reinstatement of cocaine-seeking behavior
- PMID: 16357203
- PMCID: PMC1323172
- DOI: 10.1073/pnas.0507480102
Role for hypocretin in mediating stress-induced reinstatement of cocaine-seeking behavior
Abstract
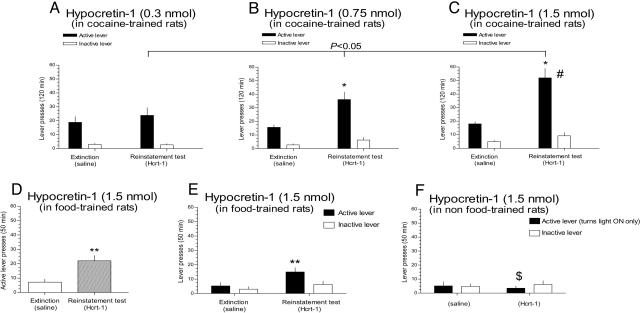
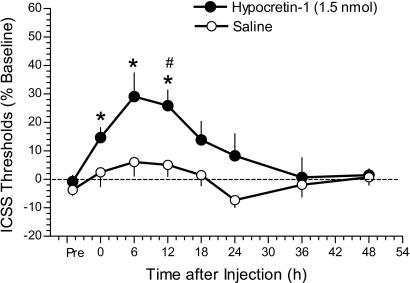
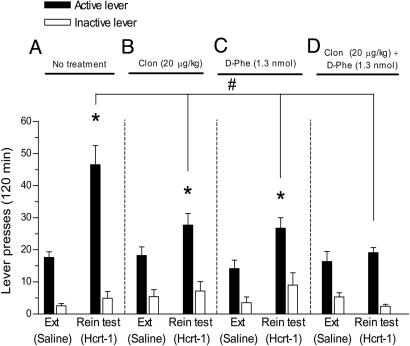
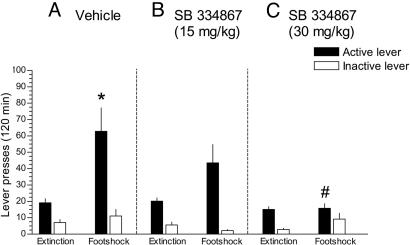
Hypocretin-1 and -2 (Hcrt-1 and Hcrt-2), also referred to as orexin-A and -B, are neuropeptides synthesized by a few thousand neurons in the lateral hypothalamus. Hypocretin-containing neurons project throughout the brain, with a prominent input to basal forebrain structures involved in motivation, reward, and stress. However, the role of hypocretins in addiction-related behaviors remains largely unexplored. Here we show that intracerebroventricular infusions of Hcrt-1 lead to a dose-related reinstatement of cocaine seeking without altering cocaine intake in rats. Hcrt-1 also dramatically elevates intracranial self-stimulation thresholds, indicating that, unlike treatments with reinforcing properties such as cocaine, Hcrt-1 negatively regulates the activity of brain reward circuitries. Hypocretin-induced reinstatement of cocaine seeking was prevented by blockade of noradrenergic and corticotropin-releasing factor systems, suggesting that Hcrt-1 reinstated drug seeking through induction of a stress-like state. Consistent with this interpretation, the selective Hcrt-1 receptor antagonist SB-334867 blocked footshock-induced reinstatement of previously extinguished cocaine-seeking behavior. These findings reveal a previously unidentified role for hypocretins in driving drug seeking through activation of stress pathways in the brain.
Figures
References
-
- O'Brien, C. P. (1997) Science 278, 66-70. - PubMed
-
- Sakurai, T., Amemiya, A., Ishii, M., Matsuzaki, I., Chemelli, R. M., Tanaka, H., Williams, S. C., Richardson, J. A., Kozlowski, G. P., Wilson, S., et al. (1998) Cell 92, 573-585. - PubMed
-
- Willie, J. T., Chemelli, R. M., Sinton, C. M. & Yanagisawa, M. (2001) Annu. Rev. Neurosci. 24, 429-458. - PubMed
-
- Taheri, S., Zeitzer, J. M. & Mignot, E. (2002) Annu. Rev. Neurosci. 25, 283-313. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
