

The Drosophila microRNA iab-4 causes a dominant homeotic transformation of halteres to wings
- PMID: 16357215
- PMCID: PMC1315399
- DOI: 10.1101/gad.1372505
The Drosophila microRNA iab-4 causes a dominant homeotic transformation of halteres to wings
Abstract
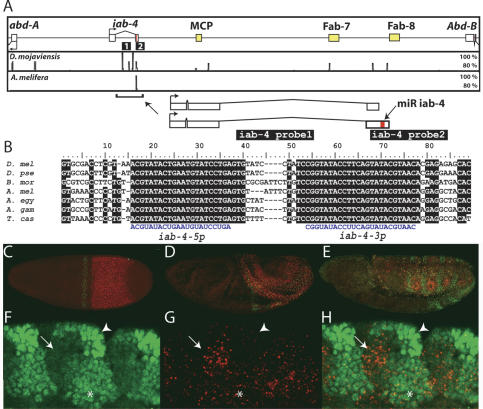
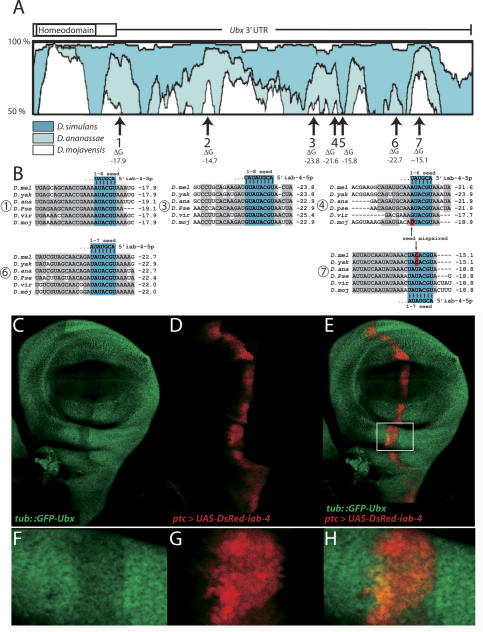
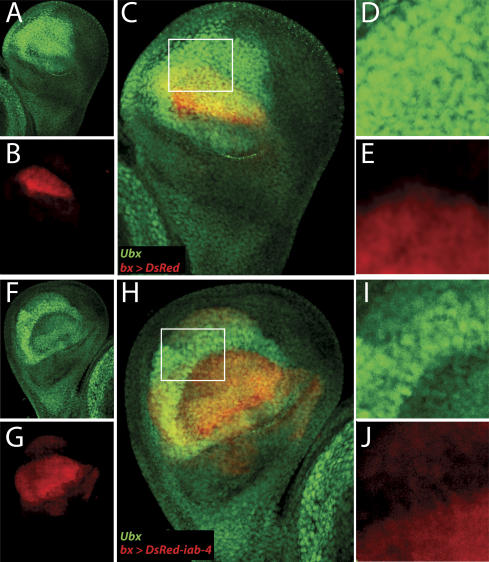
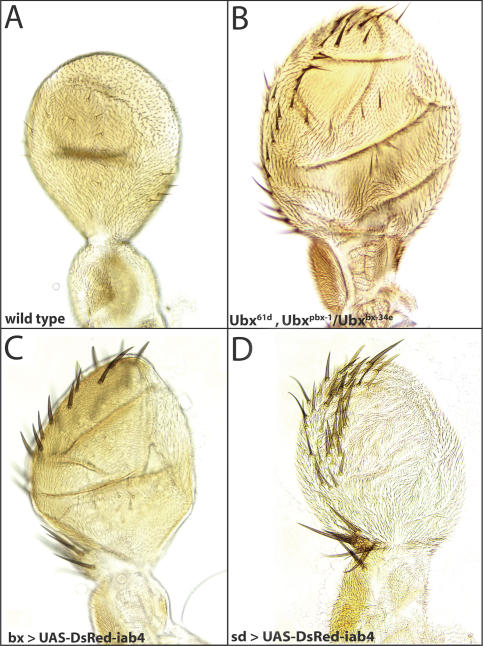
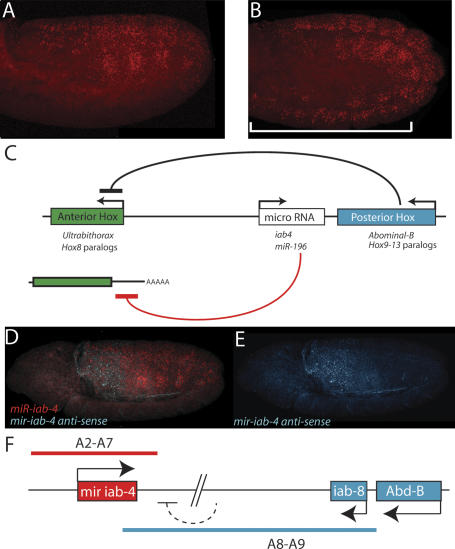
The Drosophila Bithorax Complex encodes three well-characterized homeodomain proteins that direct segment identity, as well as several noncoding RNAs of unknown function. Here, we analyze the iab-4 locus, which produces the microRNAs iab-4-5p and iab-4-3p. iab-4 is analogous to miR-196 in vertebrate Hox clusters. Previous studies demonstrate that miR-196 interacts with the Hoxb8 3' untranslated region. Evidence is presented that miR-iab-4-5p directly inhibits Ubx activity in vivo. Ectopic expression of mir-iab-4-5p attenuates endogenous Ubx protein accumulation and induces a classical homeotic mutant phenotype: the transformation of halteres into wings. These findings provide the first evidence for a noncoding homeotic gene and raise the possibility that other such genes occur within the Bithorax complex. We also discuss the regulation of mir-iab-4 expression during development.
Figures
References
-
- Aravin A.A., Lagos-Quintana, M., Yalcin, A., Zavolan, M., Marks, D., Snyder, B., Gaasterland, T., Meyer, J., and Tuschl, T. 2003. The small RNA profile during Drosophila melanogaster development. Dev. Cell 5: 337-350. - PubMed
-
- Barges S., Mihaly, J., Galloni, M., Hagstrom, K., Muller, M., Shanower, G., Schedl, P., Gyurkovics, H., and Karch, F. 2000. The Fab-8 boundary defines the distal limit of the bithorax complex iab-7 domain and insulates iab-7 from initiation elements and a PRE in the adjacent iab-8 domain. Development 127: 779-790. - PubMed
-
- Bartel D.P. 2004. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 116: 281-297. - PubMed
-
- Bentwich I., Avniel, A., Karov, Y., Aharonov, R., Gilad, S., Barad, O., Barzilai, A., Einat, P., Einav, U., Meiri, E., et al. 2005. Identification of hundreds of conserved and nonconserved human microRNAs. Nat. Genet. 37: 766-770. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases