

The core centromere and Sgo1 establish a 50-kb cohesin-protected domain around centromeres during meiosis I
- PMID: 16357219
- PMCID: PMC1315405
- DOI: 10.1101/gad.1373005
The core centromere and Sgo1 establish a 50-kb cohesin-protected domain around centromeres during meiosis I
Abstract
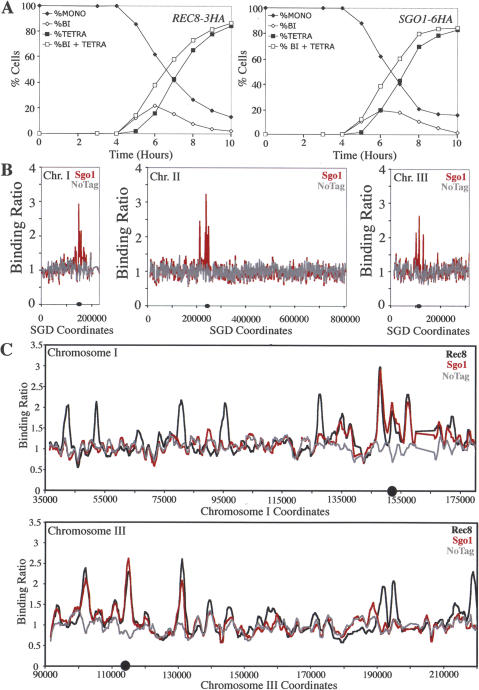
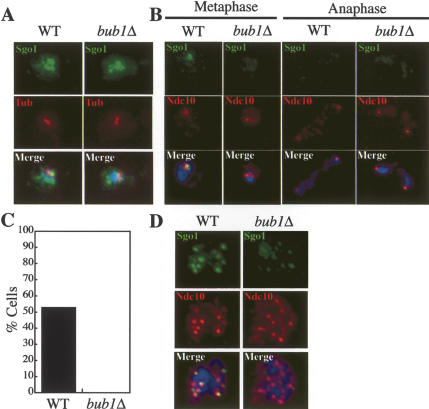
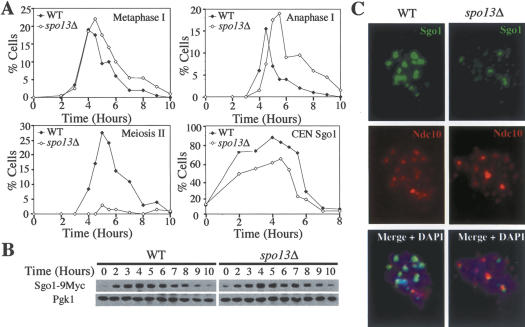
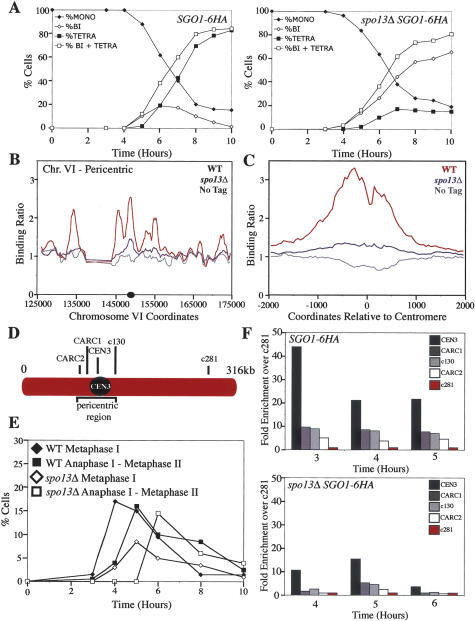
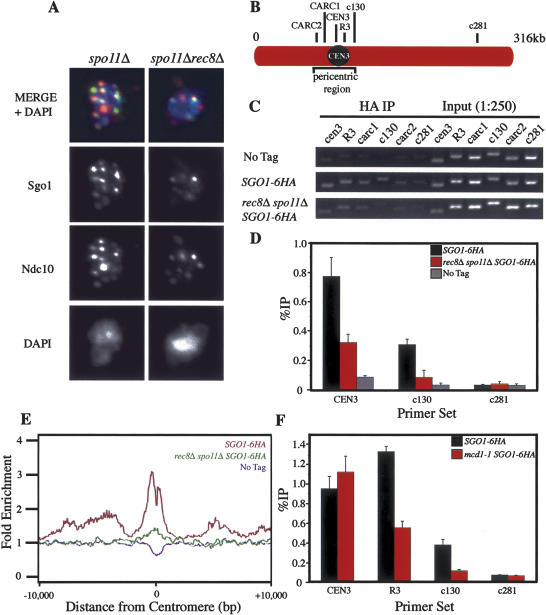
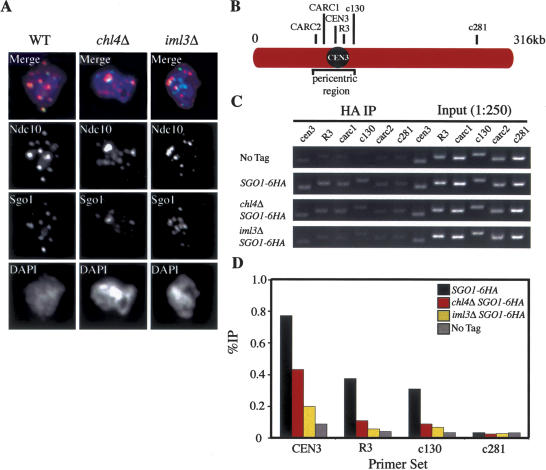
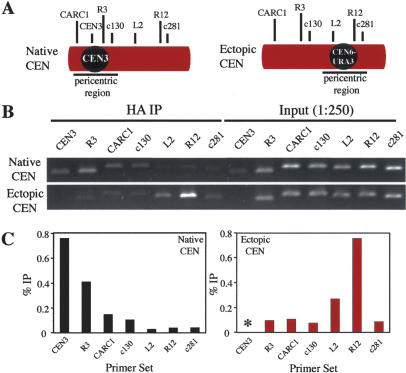
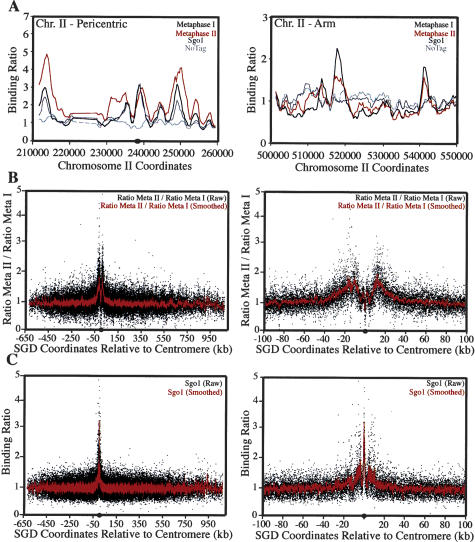
The stepwise loss of cohesins, the complexes that hold sister chromatids together, is required for faithful meiotic chromosome segregation. Cohesins are removed from chromosome arms during meiosis I but are maintained around centromeres until meiosis II. Here we show that Sgo1, a protein required for protecting centromeric cohesins from removal during meiosis I, localizes to cohesin-associated regions (CARs) at the centromere and the 50-kb region surrounding it. Establishment of this Sgo1-binding domain requires the 120-base-pair (bp) core centromere, the kinetochore component Bub1, and the meiosis-specific factor Spo13. Interestingly, cohesins and the kinetochore proteins Iml3 and Chl4 are necessary for Sgo1 to associate with pericentric regions but less so for Sgo1 to associate with the core centromeric regions. Finally, we show that the 50-kb Sgo1-binding domain is the chromosomal region where cohesins are protected from removal during meiosis I. Our results identify the portions of chromosomes where cohesins are protected from removal during meiosis I and show that kinetochore components and cohesins themselves are required to establish this cohesin protective domain.
Figures
References
-
- Aagaard L., Schmid, M., Warburton, P., and Jenuwein, T. 2000. Mitotic phosphorylation of SUV39H1, a novel component of active centromeres, coincides with transient accumulation at mammalian centromeres. J. Cell Sci. 113: 817-829. - PubMed
-
- Bernard P., Maure, J.F., and Javerzat, J.P. 2001. Fission yeast Bub1 is essential in setting up the meiotic pattern of chromosome segregation. Nat. Cell Biol. 3: 522-526. - PubMed
-
- Buonomo S.B., Clyne, R.K., Fuchs, J., Loidl, J., Uhlmann, F., and Nasmyth, K. 2000. Disjunction of homologous chromosomes in meiosis I depends on proteolytic cleavage of the meiotic cohesin Rec8 by separin. Cell 103: 387-398. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases