

TLR9 regulates Th1 responses and cooperates with TLR2 in mediating optimal resistance to Mycobacterium tuberculosis
- PMID: 16365150
- PMCID: PMC2212963
- DOI: 10.1084/jem.20051782
TLR9 regulates Th1 responses and cooperates with TLR2 in mediating optimal resistance to Mycobacterium tuberculosis
Abstract
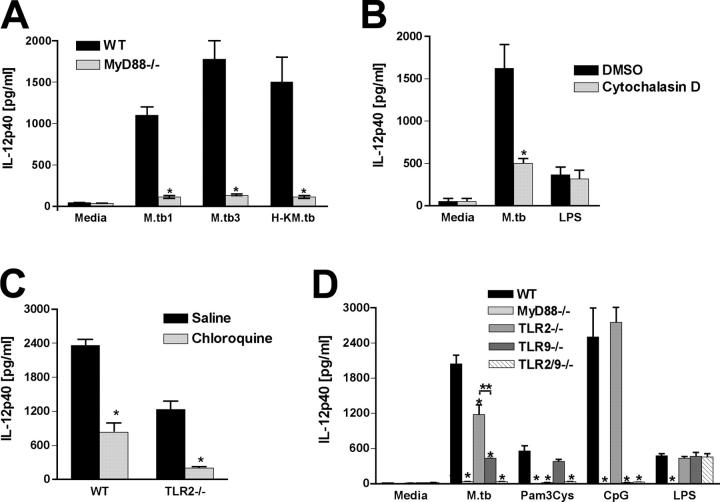
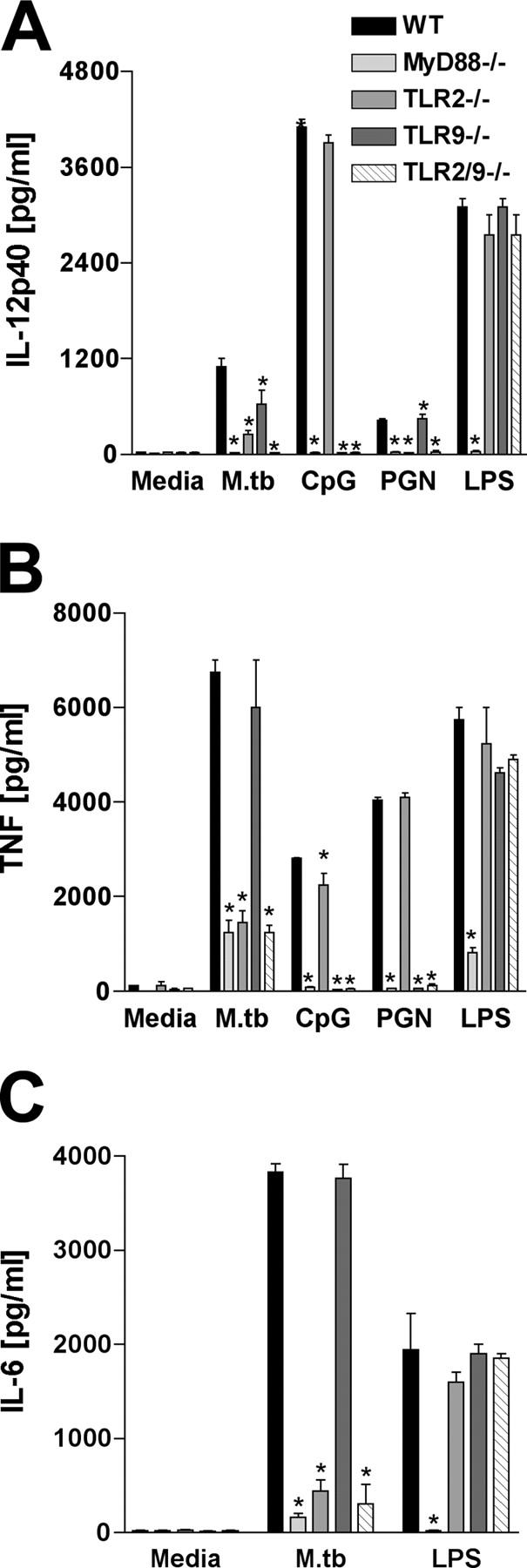
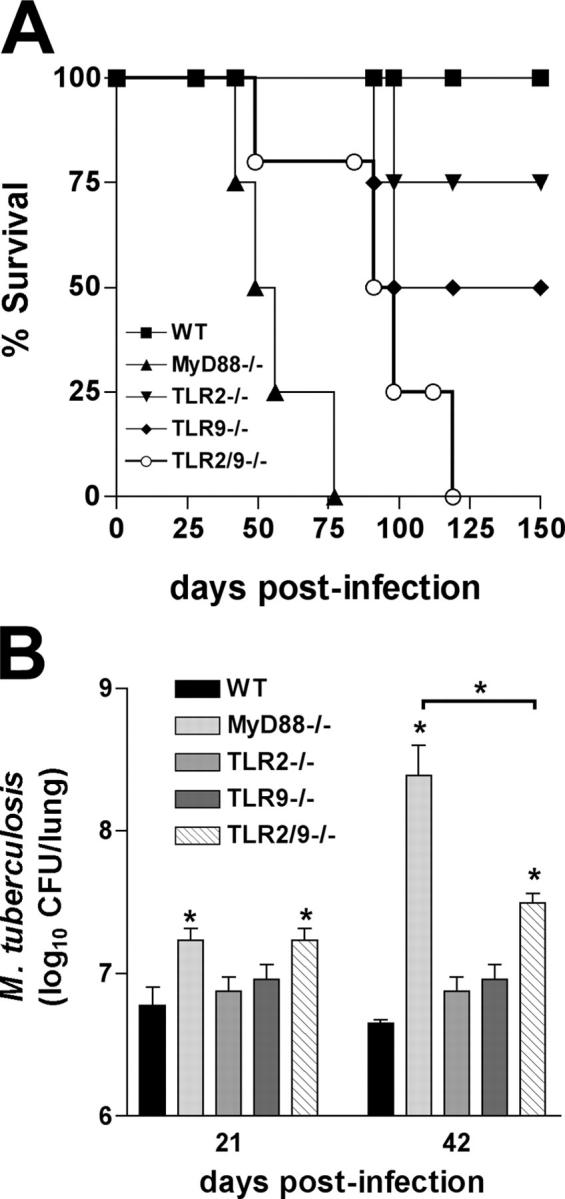
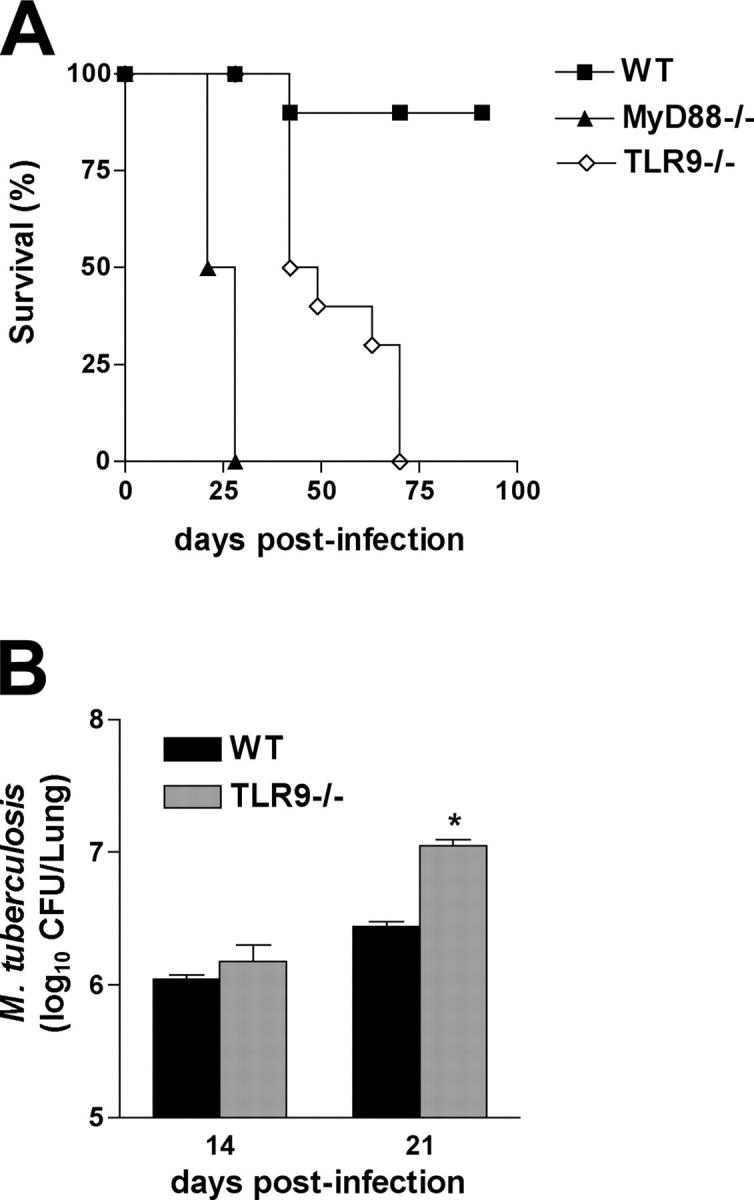
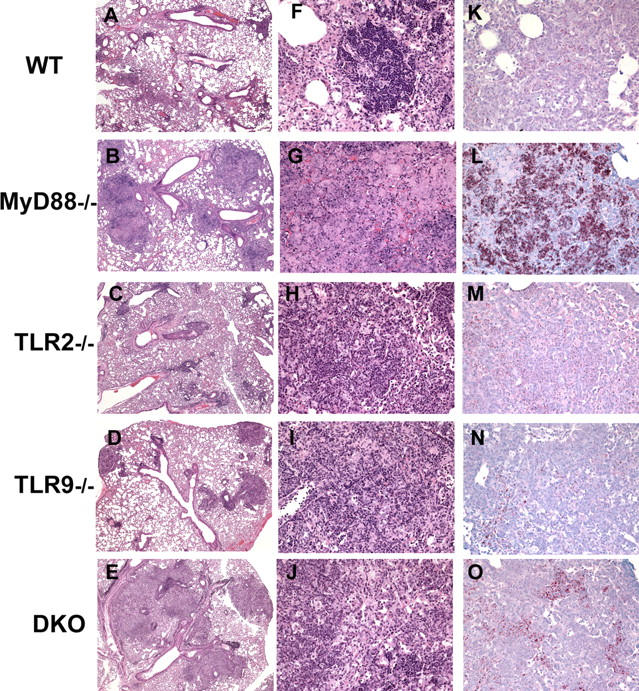
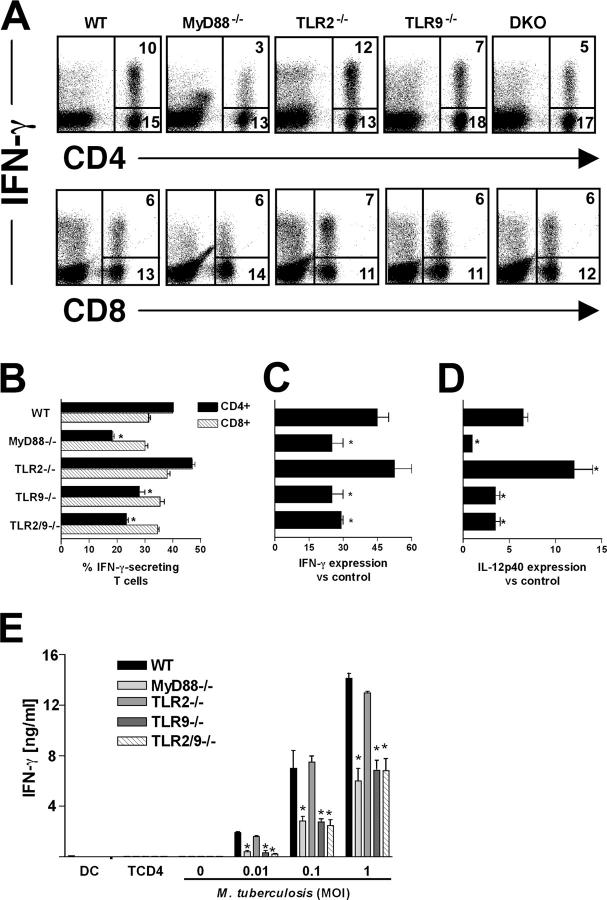
To investigate the role of Toll-like receptor (TLR)9 in the immune response to mycobacteria as well as its cooperation with TLR2, a receptor known to be triggered by several major mycobacterial ligands, we analyzed the resistance of TLR9(-/-) as well as TLR2/9 double knockout mice to aerosol infection with Mycobacterium tuberculosis. Infected TLR9(-/-) but not TLR2(-/-) mice displayed defective mycobacteria-induced interleukin (IL)-12p40 and interferon (IFN)-gamma responses in vivo, but in common with TLR2(-/-) animals, the TLR9(-/-) mice exhibited only minor reductions in acute resistance to low dose pathogen challenge. When compared with either of the single TLR-deficient animals, TLR2/9(-/-) mice displayed markedly enhanced susceptibility to infection in association with combined defects in proinflammatory cytokine production in vitro, IFN-gamma recall responses ex vivo, and altered pulmonary pathology. Cooperation between TLR9 and TLR2 was also evident at the level of the in vitro response to live M. tuberculosis, where dendritic cells and macrophages from TLR2/9(-/-) mice exhibited a greater defect in IL-12 response than the equivalent cell populations from single TLR9-deficient animals. These findings reveal a previously unappreciated role for TLR9 in the host response to M. tuberculosis and illustrate TLR collaboration in host resistance to a major human pathogen.
Figures
References
-
- Takeda, K., T. Kaisho, and S. Akira. 2003. Toll-like receptors. Annu. Rev. Immunol. 21:335–376. - PubMed
-
- Iwasaki, A., and R. Medzhitov. 2004. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 5:987–995. - PubMed
-
- Krutzik, S.R., and R.L. Modlin. 2004. The role of Toll-like receptors in combating mycobacteria. Semin. Immunol. 16:35–41. - PubMed
-
- Quesniaux, V., C. Fremond, M. Jacobs, S. Parida, D. Nicolle, V. Yeremeev, F. Bihl, F. Erard, T. Botha, M. Drennan, et al. 2004. Toll-like receptor pathways in the immune responses to mycobacteria. Microbes Infect. 6:946–959. - PubMed
-
- World Health Organization. The World Health Report 2004. Geneva. 32 pp.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
