

doi: 10.1083/jcb.200510043.
Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells
Affiliations
- PMID: 16365161
- PMCID: PMC2171318
- DOI: 10.1083/jcb.200510043
Item in Clipboard
Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells
J Cell Biol.
.
Abstract
For epithelia to function as barriers, the intercellular space must be sealed. Sealing two adjacent cells at bicellular tight junctions (bTJs) is well described with the discovery of the claudins. Yet, there are still barrier weak points at tricellular contacts, where three cells join together. In this study, we identify tricellulin, the first integral membrane protein that is concentrated at the vertically oriented TJ strands of tricellular contacts. When tricellulin expression was suppressed with RNA interference, the epithelial barrier was compromised, and tricellular contacts and bTJs were disorganized. These findings indicate the critical function of tricellulin for formation of the epithelial barrier.
Figures
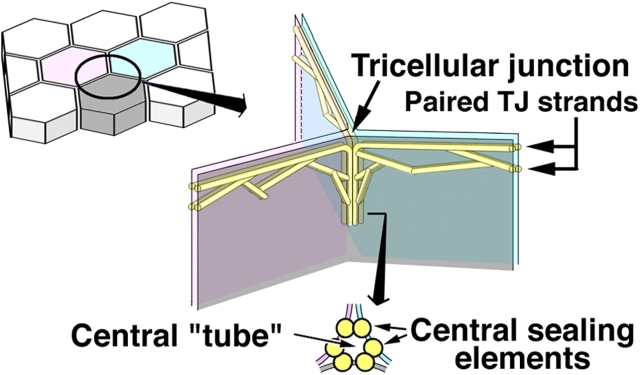
Schematic drawing of the organization of tTJs. One tricellular contact (left drawing) is enlarged in the right drawing. See the Introduction for details.
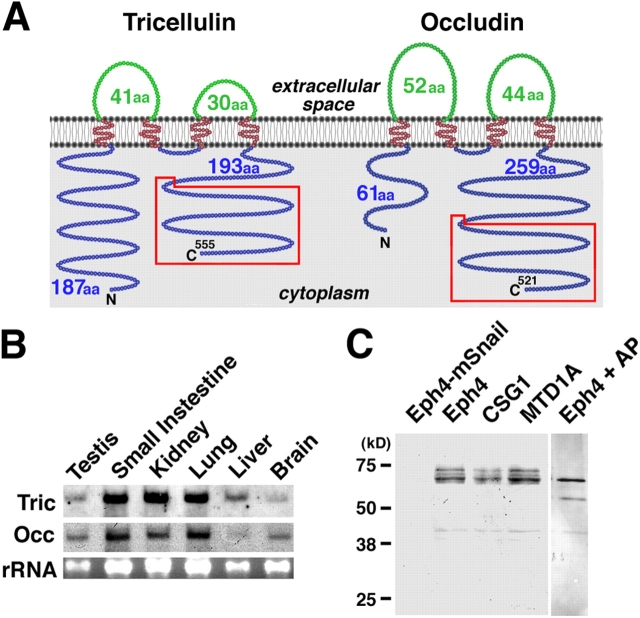
Structure and expression of mouse tricellulin. (A) Membrane folding models for mouse tricellulin and occludin. Tricellulin and occludin share two structural characteristics in addition to bearing four transmembrane domains: (1) The first extracellular loop has a high content of tyrosine and glycine residues (Fig. S1 B, available at http://www.jcb.org/cgi/content/full/jcb.200510043/DC1 ). (2) The COOH-terminal (C) ∼130 amino acids are 32% identical (boxed in red; Fig. S1 C). It should be noted that tricellulin bears a longer NH2-terminal (N) cytoplasmic domain (187 amino acids) as compared with occludin (61 amino acids). (B) Northern blotting. Both tricellulin (Tric) and occludin (Occ) are expressed in large amounts in epithelium-derived tissues. rRNA, ribosomal RNA. (C) Immunoblotting of cultured mouse epithelial cells. The total lysate of Eph4 cells exogenously expressing mouse Snail (Eph4-mSnail; Ikenouchi et al., 2003), Eph4 cells, CSG1 cells, and MTD1A cells were separated by SDS-PAGE followed by immunoblotting with anti-tricellulin pAb. Tricellulin was detected as multiple bands in Eph4, CSG1, and MTD1A cells, but after alkaline phosphatase treatment, there was only a single, smaller Mr band (Eph4 + AP). In Eph4-mSnail cells, tricellulin is undetectable.
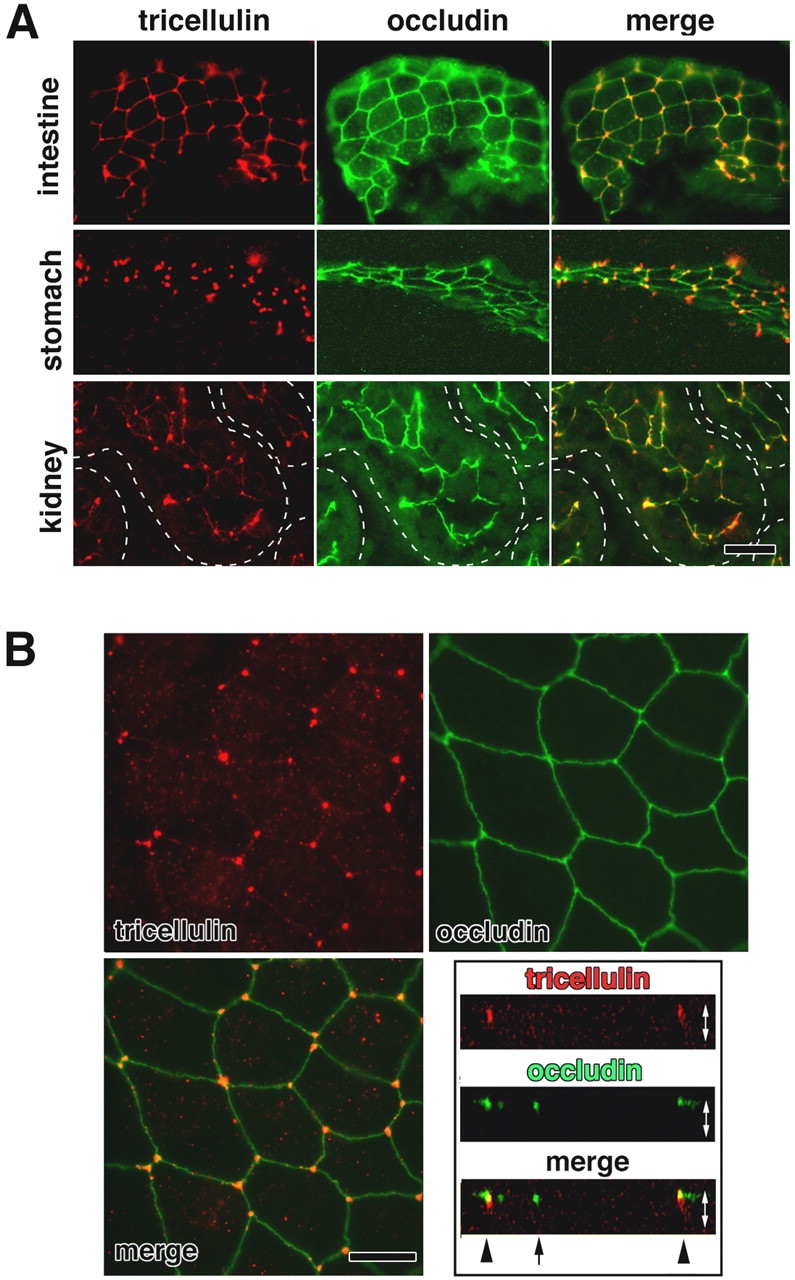
Immunofluorescence localization of tricellulin. (A) Immunofluorescence staining of frozen sections of mouse small intestine, stomach, and kidney doubly with anti-tricellulin pAb (red) and anti-occludin mAb (green). Individual renal tubules in the kidney are represented as dotted lines. Tricellulin was predominantly concentrated at the tricellular contacts, where three occludin-positive bTJs converged. There are also weaker signals at bTJs between two adjacent cells, especially in the small intestine. Bar, 20 μm. (B) Immunostaining of mouse Eph4 cells. Tricellulin (red) was concentrated at each tricellular contact, whereas occludin (green) was distributed along bTJs. The right bottom panel shows vertical sectional images generated from confocal microscopic images. Arrow and arrowheads represent bTJs and tTJs, respectively. White arrows are equivalent in length to the thickness of the cellular sheet. Bar, 10 μm.
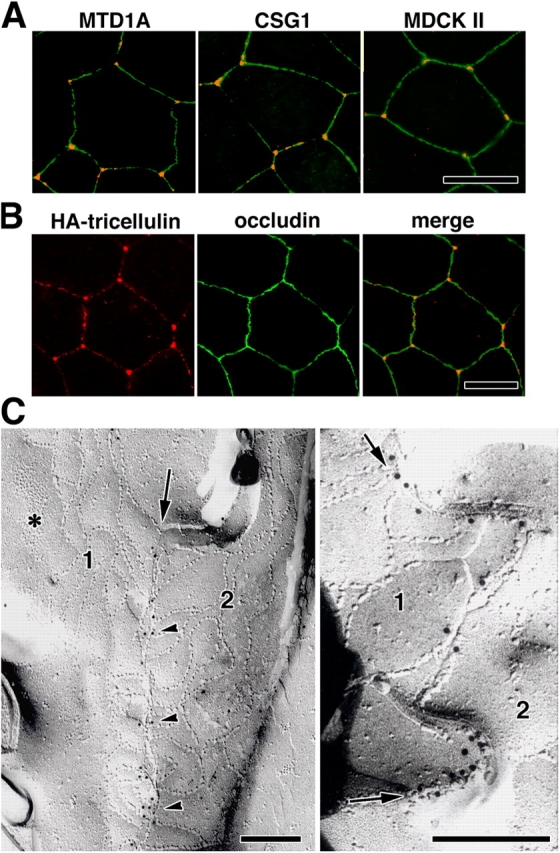
Tricellular localization of tricellulin. (A) Immunostaining of various epithelial cell lines—mouse MTD1A, mouse CSG1, and dog MDCK II cells—with anti-tricellulin pAb (red) and anti-occludin mAb (green). Tricellulin was concentrated at tricellular contacts. (B) Immunostaining of Eph4 cells expressing HA-tagged tricellulin with anti-HA mAb (red) and anti-occludin pAb (green). HA–tricellulin was recruited to the tricellular contacts in large amounts and also to occludin-positive bTJs in small amounts. (A and B) Bars, 10 μm. (C) Immunofreeze-fracture replica electron microscopy. Freeze-fracture replicas obtained from tricellular contacts of Eph4 cells were immunolabeled with anti-tricellulin pAb. In the left panel, the central sealing elements (arrow), which were identified between bTJ networks of two adjacent cells (1 and 2), were specifically labeled with anti-tricellulin pAb (arrowheads; see Fig.1). Asterisk indicates adherens junction. In the right panel, the central sealing elements between two adjacent cells (1 and 2) were enlarged, where many immunogold particles were detected (arrows). Bars, 200 nm.
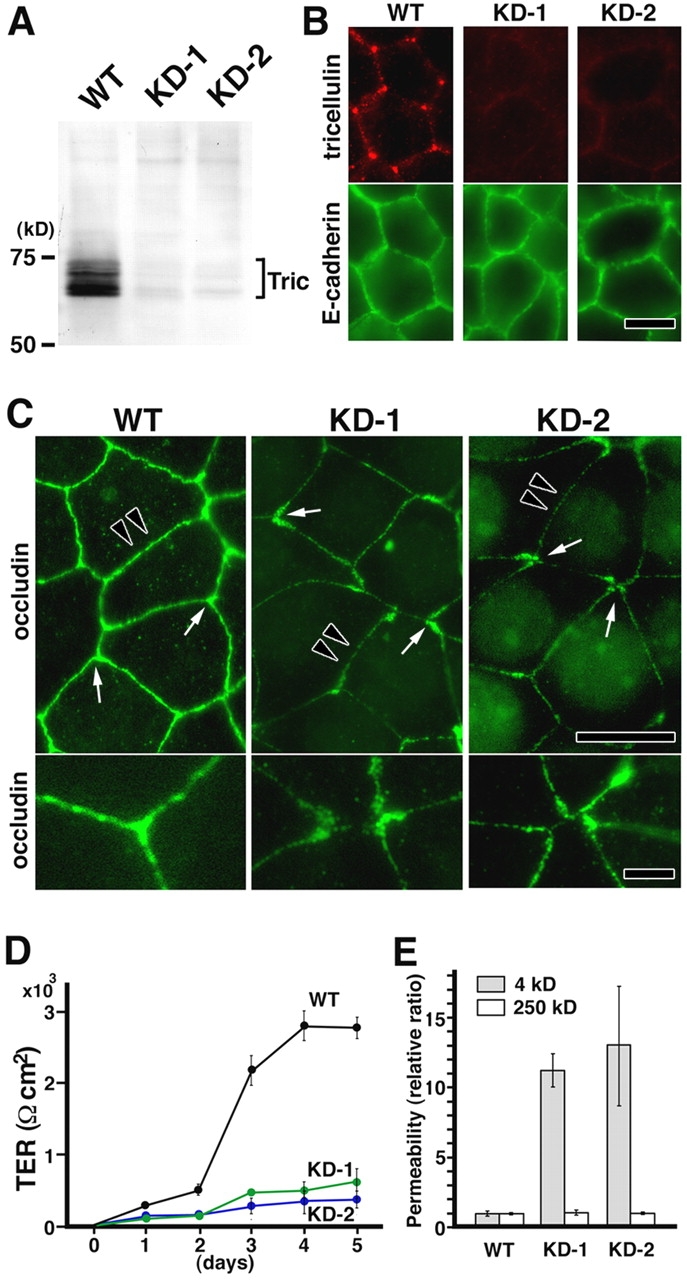
Stable suppression of tricellulin expression in Eph4 cells. (A) Immunoblot analysis using anti-tricellulin pAb. Two independent Eph4 cell clones with suppressed tricellulin expression (KD-1 and KD-2) were established by stably expressing two distinct short interfering RNAs (Brummelkamp et al., 2002). Equal amounts of total proteins were analyzed by SDS-PAGE and immunoblotting. Tric, tricellulin. (B) Immunofluorescence staining of wild-type (WT) and tricellulin RNAi knockdown (KD-1 and KD-2) cells with anti-tricellulin pAb (red) and anti–E-cadherin mAb (green). Bar, 10 μm. (C) Immunofluorescence staining of WT, KD-1, and KD-2 cells with anti-occludin mAb. In KD-1 and KD-2 cells, both ends of occludin-positive bTJs showed characteristic teardrop-like shapes, and the tricellular contacts were disorganized with gaps (arrows in top panels; enlarged in bottom panels). Furthermore, in these cells, the bTJs were significantly thinner than those in wild-type Eph4 cells (arrowheads in top panels). Bars (top), 10 μm; (bottom) 3 μm. (D) TER measurements of WT, KD-1, and KD-2 cells. Tricellulin knockdown significantly affected the development of the epithelial barrier (n = 10 for each cell line). (E) FITC–dextran flux measurements of WT, KD-1, and KD-2 cells. There was a significant difference in the flux of 4-kD (but not 250 kD) FITC–dextran across monolayers between wild-type Eph4 cells and KD-1/2 cells (n = 10 for each cell line).
References
-
- Anderson, J.M., C.M. Van Itallie, and A.S. Fanning. 2004. Setting up a selective barrier at the apical junction complex. Curr. Opin. Cell Biol. 16:140–145. - PubMed
-
- Brummelkamp, T.R., R. Bernards, and R. Agami. 2002. A system for stable expression of short interfering RNAs in mammalian cells. Science. 296:550–553. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
