

L1 integration in a transgenic mouse model
- PMID: 16365384
- PMCID: PMC1361720
- DOI: 10.1101/gr.4571606
L1 integration in a transgenic mouse model
Abstract
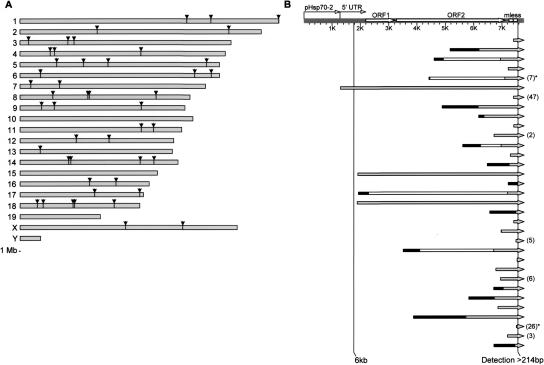
To study integration of the human LINE-1 retrotransposon (L1) in vivo, we developed a transgenic mouse model of L1 retrotransposition that displays de novo somatic L1 insertions at a high frequency, occasionally several insertions per mouse. We mapped 3' integration sites of 51 insertions by Thermal Asymmetric Interlaced PCR (TAIL-PCR). Analysis of integration locations revealed a broad genomic distribution with a modest preference for intergenic regions. We characterized the complete structures of 33 de novo retrotransposition events. Our results highlight the large number of highly truncated L1s, as over 52% (27/51) of total integrants were <1/3 the length of a full-length element. New integrants carry all structural characteristics typical of genomic L1s, including a number with inversions, deletions, and 5'-end microhomologies to the target DNA sequence. Notably, at least 13% (7/51) of all insertions contain a short stretch of extra nucleotides at their 5' end, which we postulate result from template-jumping by the L1-encoded reverse transcriptase. We propose a unified model of L1 integration that explains all of the characteristic features of L1 retrotransposition, such as 5' truncations, inversions, extra nucleotide additions, and 5' boundary and inversion point microhomologies.
Figures




References
-
- Bibillo, A. and Eickbush, T.H. 2002a. High processivity of the reverse transcriptase from a non-long terminal repeat retrotransposon. J. Biol. Chem. 277 34836-34845. - PubMed
-
- ———. 2002b. The reverse transcriptase of the R2 non-LTR retrotransposon: Continuous synthesis of cDNA on non-continuous RNA templates. J. Mol. Biol. 316 459-473. - PubMed
-
- ———. 2004. End-to-end template jumping by the reverse transcriptase encoded by the R2 retrotransposon. J. Biol. Chem. 279 14945-14953. - PubMed
-
- Boissinot, S., Chevret, P., and Furano, A.V. 2000. L1 (LINE-1) retrotransposon evolution and amplification in recent human history. Mol. Biol. Evol. 17 915-928. - PubMed
-
- Boissinot, S., Entezam, A., and Furano, A.V. 2001. Selection against deleterious LINE-1-containing loci in the human lineage. Mol. Biol. Evol. 18 926-935. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases