

Dissection of ESAT-6 system 1 of Mycobacterium tuberculosis and impact on immunogenicity and virulence
- PMID: 16368961
- PMCID: PMC1346617
- DOI: 10.1128/IAI.74.1.88-98.2006
Dissection of ESAT-6 system 1 of Mycobacterium tuberculosis and impact on immunogenicity and virulence
Abstract
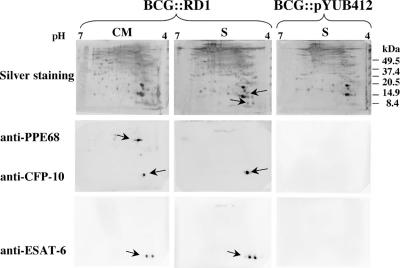
The dedicated secretion system ESX-1 of Mycobacterium tuberculosis encoded by the extended RD1 region (extRD1) assures export of the ESAT-6 protein and its partner, the 10-kDa culture filtrate protein CFP-10, and is missing from the vaccine strains M. bovis BCG and M. microti. Here, we systematically investigated the involvement of each individual ESX-1 gene in the secretion of both antigens, specific immunogenicity, and virulence. ESX-1-complemented BCG and M. microti strains were more efficiently engulfed by bone-marrow-derived macrophages than controls, and this may account for the enhanced in vivo growth of ESX-1-carrying strains. Inactivation of gene pe35 (Rv3872) impaired expression of CFP-10 and ESAT-6, suggesting a role in regulation. Genes Rv3868, Rv3869, Rv3870, Rv3871, and Rv3877 encoding an ATP-dependent chaperone and translocon were essential for secretion of ESAT-6 and CFP-10 in contrast to ppe68 Rv3873 and Rv3876, whose inactivation did not impair secretion of ESAT-6. A strict correlation was found between ESAT-6 export and the generation of ESAT-6 specific T-cell responses in mice. Furthermore, ESAT-6 secretion and specific immunogenicity were almost always correlated with enhanced virulence in the SCID mouse model. Only loss of Rv3865 and part of Rv3866 did not affect ESAT-6 secretion or immunogenicity but led to attenuation. This suggests that Rv3865/66 represent a new virulence factor that is independent from ESAT-6 secretion. The present study has allowed us to identify new aspects of the extRD1 region of M. tuberculosis and to explore its role in the pathogenesis of tuberculosis.
Figures




References
-
- Bahk, Y. Y., S. A. Kim, J. S. Kim, H. J. Euh, G. H. Bai, S. N. Cho, and Y. S. Kim. 2004. Antigens secreted from Mycobacterium tuberculosis: identification by proteomics approach and test for diagnostic marker. Proteomics 4:3299-3307. - PubMed
-
- Bange, F. C., F. M. Collins, and W. R. Jacobs, Jr. 1999. Survival of mice infected with Mycobacterium smegmatis containing large DNA fragments from Mycobacterium tuberculosis. Tuberc. Lung Dis. 79:171-180. - PubMed
-
- Behr, M. A., M. A. Wilson, W. P. Gill, H. Salamon, G. K. Schoolnik, S. Rane, and P. M. Small. 1999. Comparative genomics of BCG vaccines by whole-genome DNA microarray. Science 284:1520-1523. - PubMed
-
- Berthet, F. X., P. B. Rasmussen, I. Rosenkrands, P. Andersen, and B. Gicquel. 1998. A Mycobacterium tuberculosis operon encoding ESAT-6 and a novel low-molecular-mass culture filtrate protein (CFP-10). Microbiology 144:3195-3203. - PubMed
-
- Braunstein, M., B. J. Espinosa, J. Chan, J. T. Belisle, and W. R. Jacobs, Jr. 2003. SecA2 functions in the secretion of superoxide dismutase A and in the virulence of Mycobacterium tuberculosis. Mol. Microbiol. 48:453-464. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
