

Barrier-to-autointegration factor phosphorylation on Ser-4 regulates emerin binding to lamin A in vitro and emerin localization in vivo
- PMID: 16371512
- PMCID: PMC1382305
- DOI: 10.1091/mbc.e05-04-0356
Barrier-to-autointegration factor phosphorylation on Ser-4 regulates emerin binding to lamin A in vitro and emerin localization in vivo
Abstract
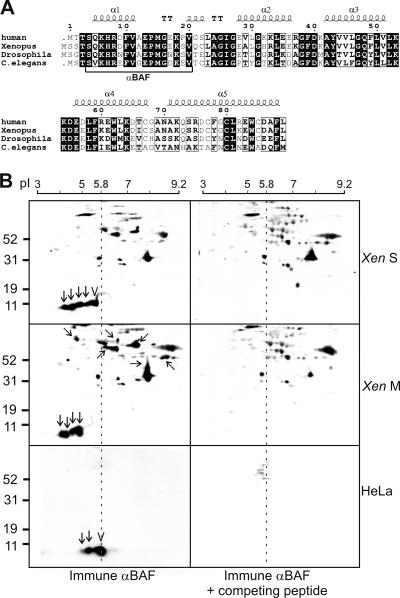
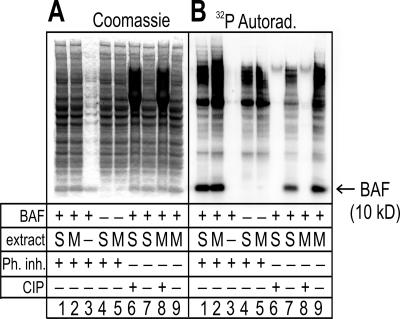
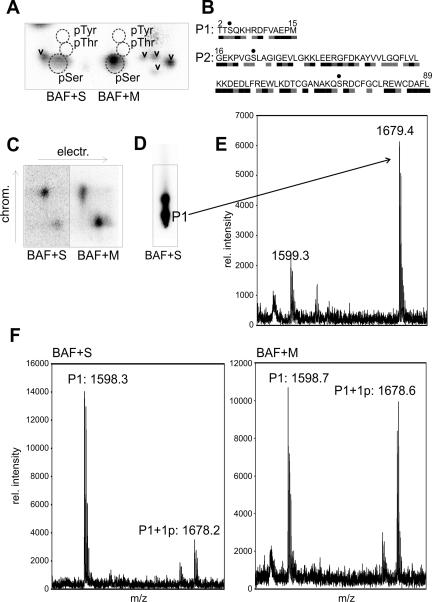
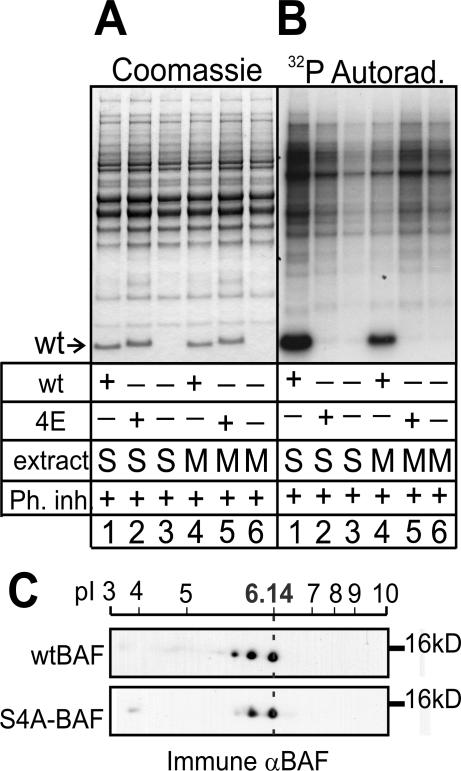
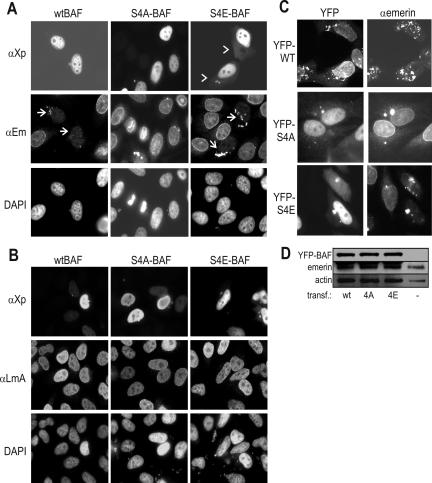
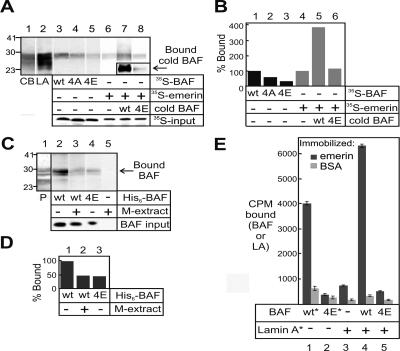
Barrier-to-autointegration factor (BAF) is a conserved 10-kDa chromatin protein essential in proliferating cells. BAF dimers bind double-stranded DNA, histone H3, histone H1.1, lamin A, and transcription regulators, plus emerin and other LEM-domain nuclear proteins. Two-dimensional gel analysis showed that endogenous human and Xenopus BAF are posttranslationally modified by phosphorylation and potentially other modifications and that they are hyperphosphorylated during mitosis. The invariant Ser-4 residue on BAF is a major site of phosphorylation during both interphase and mitosis. In HeLa cells that overexpressed the phosphomimetic BAF missense mutant S4E, but not S4A, emerin mislocalized from the nuclear envelope, suggesting Ser-4-nonphosphorylated BAF normally promotes emerin localization at the nuclear envelope. Supporting this model, wild-type BAF but not mutant S4E enhanced emerin binding to lamin A in vitro. Thus, Ser-4-unphosphorylated BAF has a positive role in localizing emerin; this role may be disease relevant because loss or mislocalization of emerin causes Emery-Dreifuss muscular dystrophy. Our findings further suggest Ser-4 phosphorylation inhibits BAF binding to emerin and lamin A, and thereby weakens emerin-lamin interactions during both mitosis and interphase.
Figures
References
-
- Bengtsson, L., and Wilson, K. L. (2004). Multiple and surprising new functions for emerin, a nuclear membrane protein. Curr. Opin. Cell Biol. 16, 73–79. - PubMed
-
- Bradley, C. M., Ronning, D. R., Ghirlando, R., Craigie, R., and Dyda, F. (2005). Structural basis for DNA bridging by barrier-to-autointegration factor. Nat. Struct. Mol. Biol. 12, 935–936. - PubMed
-
- Broers, J. L., Hutchison, C. J., and Ramaekers, F. C. (2004). Laminopathies. J. Pathol. 204, 478–488. - PubMed
-
- Cai, M., Huang, Y., Zheng, R., Wei, S. Q., Ghirlando, R., Lee, M. S., Craigie, R., Gronenborn, A. M., and Clore, G. M. (1998). Solution structure of the cellular factor BAF responsible for protecting retroviral DNA from autointegration. Nat. Struct. Biol. 5, 903–909. - PubMed
-
- Casaday, R. J., Bailey, J. R., Kalb, S. R., Brignole, E. J., Loveland, A. N., Cotter, R. J., and Gibson, W. (2004). Assembly protein precursor (pU.L80.5 homolog) of simian cytomegalovirus is phosphorylated at a glycogen synthase kinase 3 site and its downstream “priming” site: phosphorylation affects interactions of protein with itself and with major capsid protein. J. Virol. 78, 13501–13511. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
