

Calcium at fertilization and in early development
- PMID: 16371595
- PMCID: PMC3299562
- DOI: 10.1152/physrev.00023.2005
Calcium at fertilization and in early development
Abstract
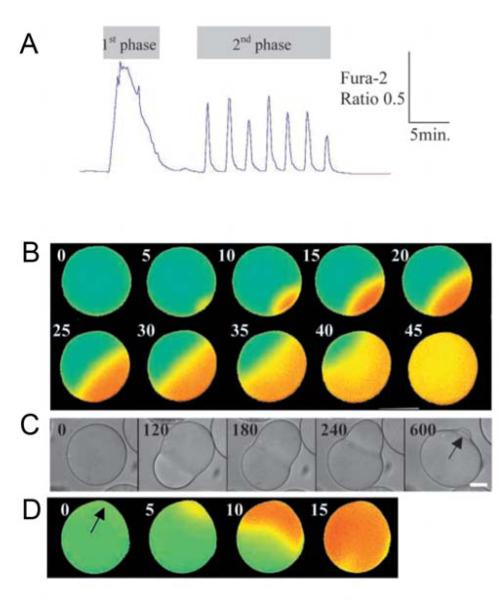
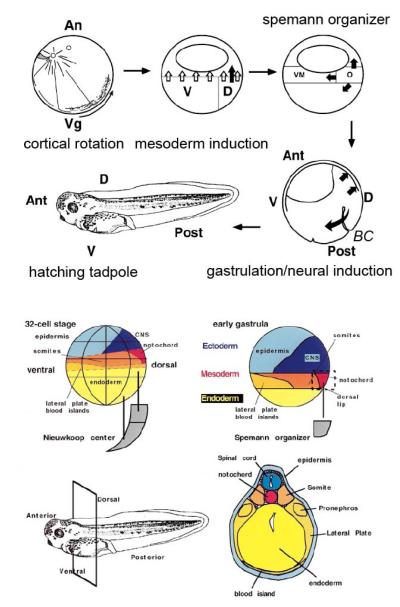
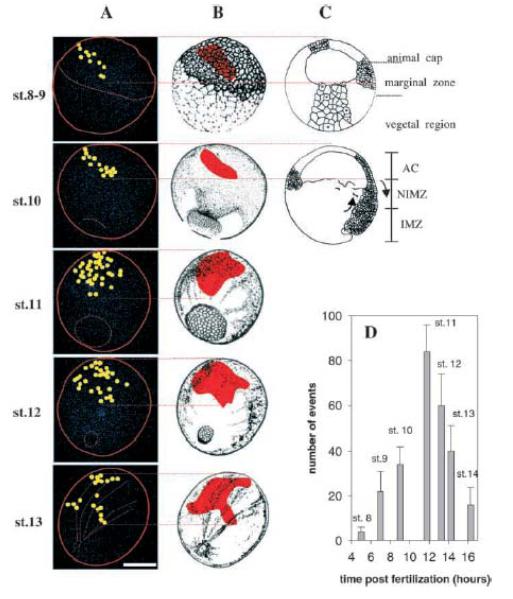
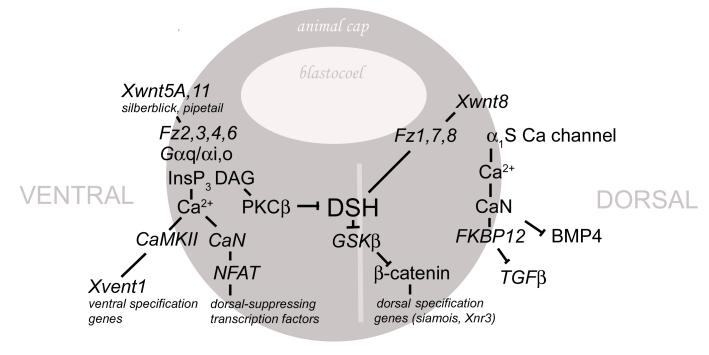
Fertilization calcium waves are introduced, and the evidence from which we can infer general mechanisms of these waves is presented. The two main classes of hypotheses put forward to explain the generation of the fertilization calcium wave are set out, and it is concluded that initiation of the fertilization calcium wave can be most generally explained in invertebrates by a mechanism in which an activating substance enters the egg from the sperm on sperm-egg fusion, activating the egg by stimulating phospholipase C activation through a src family kinase pathway and in mammals by the diffusion of a sperm-specific phospholipase C from sperm to egg on sperm-egg fusion. The fertilization calcium wave is then set into the context of cell cycle control, and the mechanism of repetitive calcium spiking in mammalian eggs is investigated. Evidence that calcium signals control cell division in early embryos is reviewed, and it is concluded that calcium signals are essential at all three stages of cell division in early embryos. Evidence that phosphoinositide signaling pathways control the resumption of meiosis during oocyte maturation is considered. It is concluded on balance that the evidence points to a need for phosphoinositide/calcium signaling during resumption of meiosis. Changes to the calcium signaling machinery occur during meiosis to enable the production of a calcium wave in the mature oocyte when it is fertilized; evidence that the shape and structure of the endoplasmic reticulum alters dynamically during maturation and after fertilization is reviewed, and the link between ER dynamics and the cytoskeleton is discussed. There is evidence that calcium signaling plays a key part in the development of patterning in early embryos. Morphogenesis in ascidian, frog, and zebrafish embryos is briefly described to provide the developmental context in which calcium signals act. Intracellular calcium waves that may play a role in axis formation in ascidian are discussed. Evidence that the Wingless/calcium signaling pathway is a strong ventralizing signal in Xenopus, mediated by phosphoinositide signaling, is adumbrated. The central role that calcium channels play in morphogenetic movements during gastrulation and in ectodermal and mesodermal gene expression during late gastrulation is demonstrated. Experiments in zebrafish provide a strong indication that calcium signals are essential for pattern formation and organogenesis.
Figures
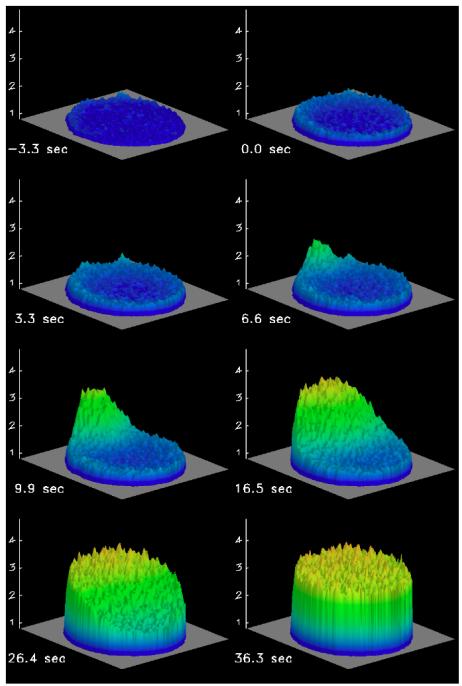
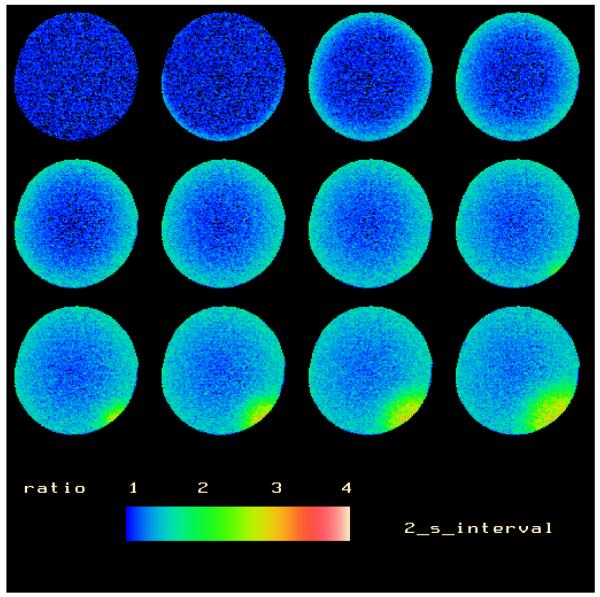
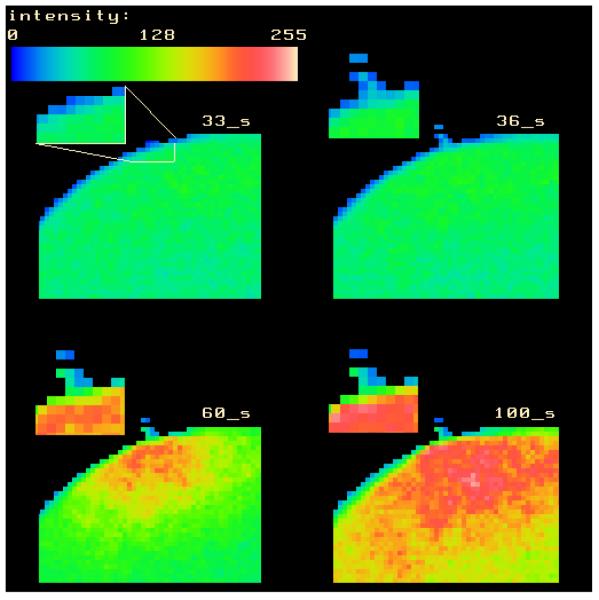

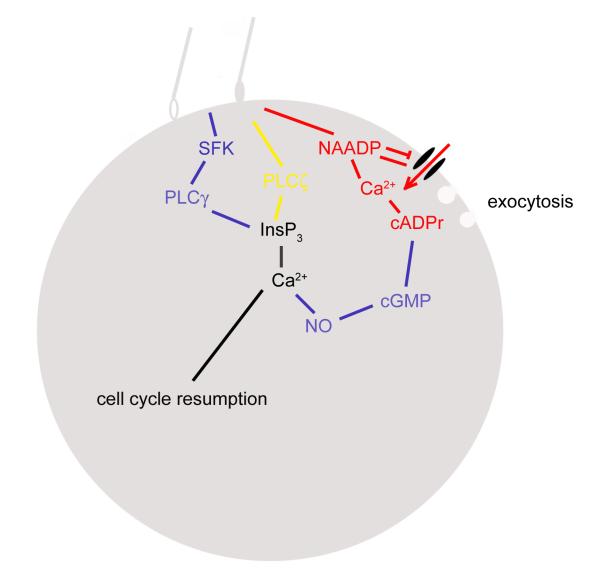

References
-
- Aanstad P, Whitaker M. Predictability of dorso-ventral asymmetry in the cleavage stage zebrafish embryo: an analysis using lithium sensitivity as a dorso-ventral marker. Mech Dev. 1999;88:33–41. - PubMed
-
- Aarhus R, Dickey DM, Graeff RM, Gee KR, Walseth TF, Lee HC. Activation and inactivation of Ca2+ release by NAADP+ J Biol Chem. 1996;271:8513–8516. - PubMed
-
- Aarhus R, Graeff RM, Dickey DM, Walseth TF, Lee HC. ADP-ribosyl cyclase and CD38 catalyze the synthesis of a calcium-mobilizing metabolite from NADP. 1995;270:30327–30333. - PubMed
-
- Abassi YA, Carroll DJ, Giusti AF, Belton RJ, Foltz KR. Evidence that Src-type tyrosine kinase activity is necessary for initiation of calcium release at fertilization in sea urchin eggs. Dev Biol. 2000;218:206–219. - PubMed
-
- Abassi YA, Foltz KR. Tyrosine phosphorylation of the egg receptor for sperm at fertilization. Dev Biol. 1994;164:430–443. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous