

Anther cap retention prevents self-pollination by elaterid beetles in the South African orchid Eulophia foliosa
- PMID: 16373371
- PMCID: PMC2803648
- DOI: 10.1093/aob/mcj041
Anther cap retention prevents self-pollination by elaterid beetles in the South African orchid Eulophia foliosa
Abstract
Background and aims: Pollination by insects that spend long periods visiting many flowers on a plant may impose a higher risk of facilitated self-pollination. Orchids and asclepiads are particularly at risk as their pollen is packaged as pollinia and so can be deposited on self-stigmas en masse. Many orchids and asclepiads have adaptations to limit self-deposition of pollinia, including gradual reconfiguration of pollinaria following removal. Here an unusual mechanism--anther cap retention--that appears to prevent self-pollination in the South African orchid Eulophia foliosa is examined.
Methods: Visits to inflorescences in the field were observed and pollinators collected. Visitation rates to transplanted inflorescences were compared between a site where putative pollinators were abundant and a site where they were rare. Anther cap retention times were determined for removed pollinaria and atmospheric vapour pressure deficit was recorded concurrently. Anther cap anatomy was examined using light microscopy.
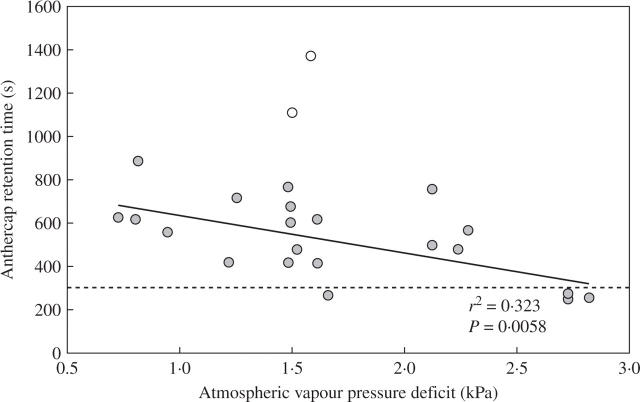
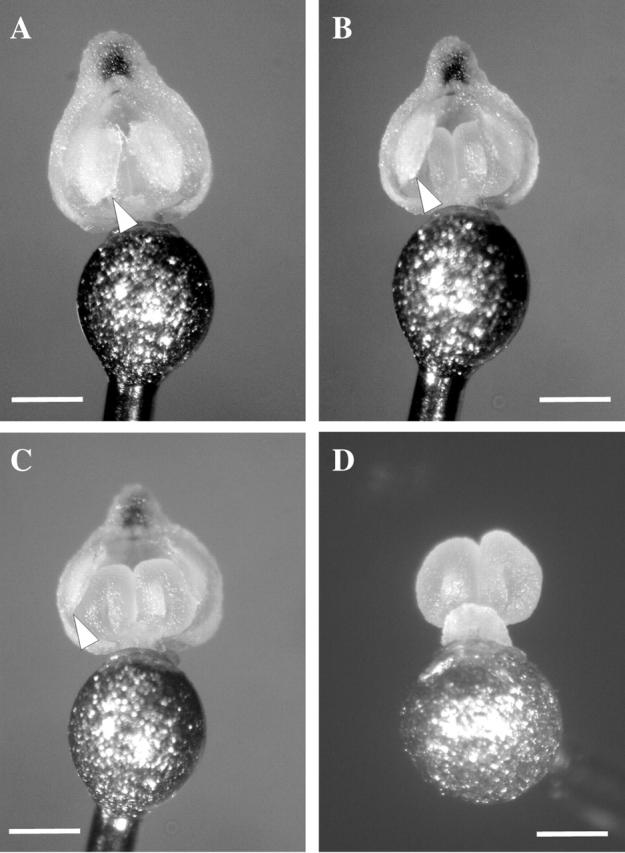
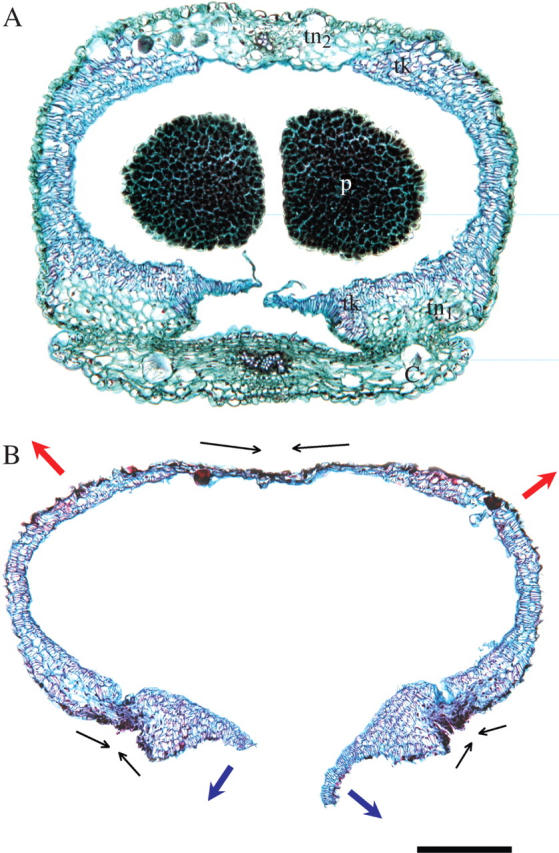
Key results: Eulophia foliosa is pollinated almost exclusively by Cardiophorus obliquemaculatus (Elateridae) beetles, which remain on the deceptive inflorescences for on average 301 s (n = 18). The anther cap that covers the pollinarium is retained for an average of 512 s (n = 24) after pollinarium removal by beetles. In all populations measured, anther cap dimensions are greater than those of the stigmatic cavity, thus precluding the deposition of self-pollinia until after the anther cap has dropped. An anatomical investigation of this mechanism suggests that differential water loss from regions of the anther cap results in opening of the anther cap flaps. This is supported by observations that as atmospheric vapour pressure deficits increased, the duration of anther cap retention was reduced.
Conclusions: Flowers of E. foliosa are specialized for pollination by elaterid beetles. Retention of anther caps for a period exceeding average visit times by beetles to inflorescences appears to prevent facilitated self-pollination in E. foliosa effectively.
Figures

References
-
- Barrett SCH. 2002. Sexual interference of the floral kind. Heredity 88: 154–159. - PubMed
-
- Bernhardt P. 2000. Convergent evolution and adaptive radiation of beetle-pollinated angiosperms. Plant Systematics and Evolution 222: 293–320.
-
- Bookman SS. 1981. The floral morphology of Asclepias speciosa (Asclepiadaceae) in relation to pollination and a clarification in terminology for the genus. American Journal of Botany 68: 675–679.
-
- Borba EL, Semir J. 1999. Temporal variation in pollinarium size after its removal in species of Bulbophyllum: a different mechanism preventing self-pollination in Orchidaceae. Plant Systematics and Evolution 217: 197–204.
-
- Borba EL, Semir J. 2001. Pollinator specificity and convergence in fly-pollinated Pleurothallis (Orchidaceae) species: a multiple population approach. Annals of Botany 88: 75–88.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
