

Differential polyamine sensitivity in inwardly rectifying Kir2 potassium channels
- PMID: 16373386
- PMCID: PMC1796793
- DOI: 10.1113/jphysiol.2005.097741
Differential polyamine sensitivity in inwardly rectifying Kir2 potassium channels
Abstract
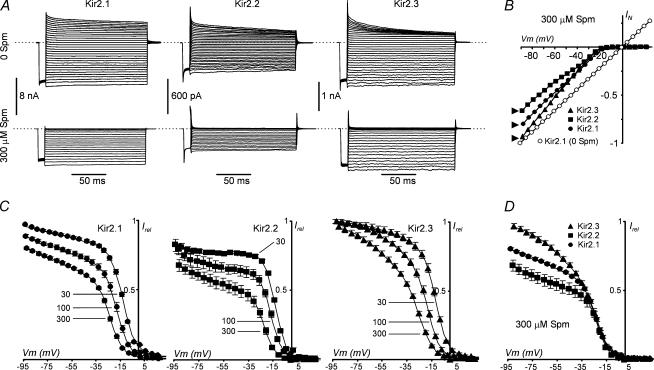
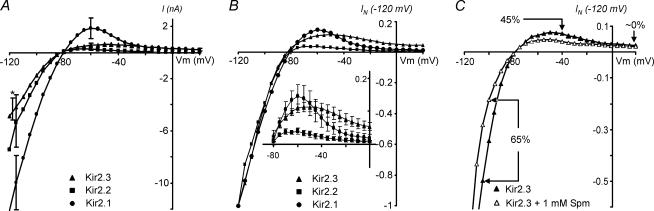
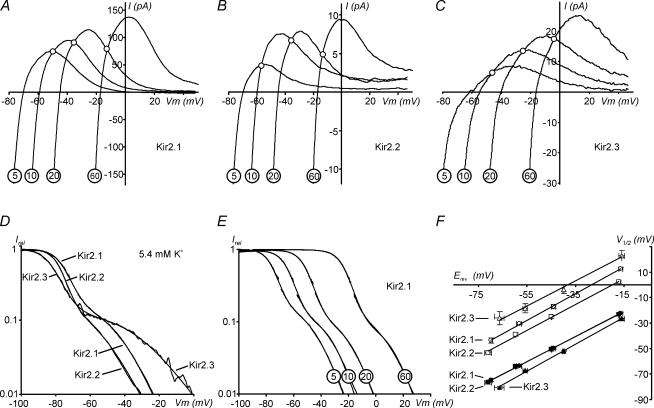
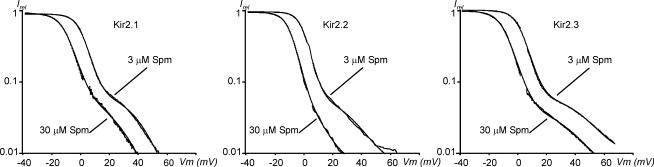
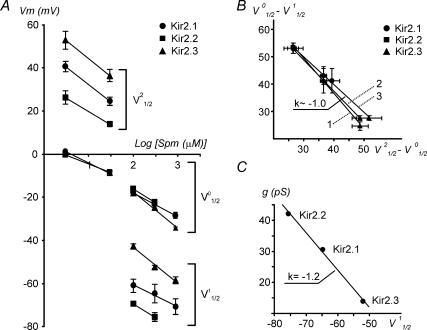
Recent studies have shown that Kir2 channels display differential sensitivity to intracellular polyamines, and have raised a number of questions about several properties of inward rectification important to the understanding of their physiological roles. In this study, we have carried out a detailed characterization of steady-state and kinetic properties of block of Kir2.1-3 channels by spermine. High-resolution recordings from outside-out patches showed that in all Kir2 channels current-voltage relationships display a 'crossover' effect upon change in extracellular K+. Experiments at different concentrations of spermine allowed for the characterization of two distinct shallow components of rectification, with the voltages for half-block negative (V1(1/2)) and positive (V2(1/2)) to the voltage of half-block for the major steep component of rectification (V0(1/2)). While V1(1/2) and V2(1/2) voltages differ significantly between Kir2 channels, they were coupled to each other according to the equation V1(1/2)-V2(1/2) = constant, strongly suggesting that similar structures may underlie both components. In Kir2.3 channels, the V2(1/2) was approximately 50 mV positive to V0(1/2), leading to a pattern of outward currents distinct from that of Kir2.1 and Kir2.2 channels. The effective valency of spermine block (Z0) was highest in Kir2.2 channels while the valencies in Kir2.1 and Kir2.3 channels were not significantly different. The voltage dependence of spermine unblock was similar in all Kir2 channels, but the rates of unblock were approximately 7-fold and approximately 16-fold slower in Kir2.3 channels than those in Kir2.1 and Kir2.2 when measured at high and physiological extracellular K+, respectively. In all Kir2 channels, the instantaneous phase of activation was present. The instantaneous phase was difficult to resolve at high extracellular K+ but it became evident and accounted for nearly 30-50% of the total current when recorded at physiological extracellular K+. In conclusion, the data are consistent with the universal mechanism of rectification in Kir2 channels, but also point to significant, and physiologically important, quantitative differences between Kir2 isoforms.
Figures
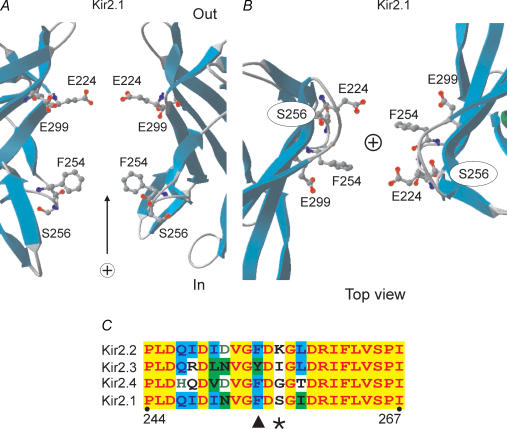
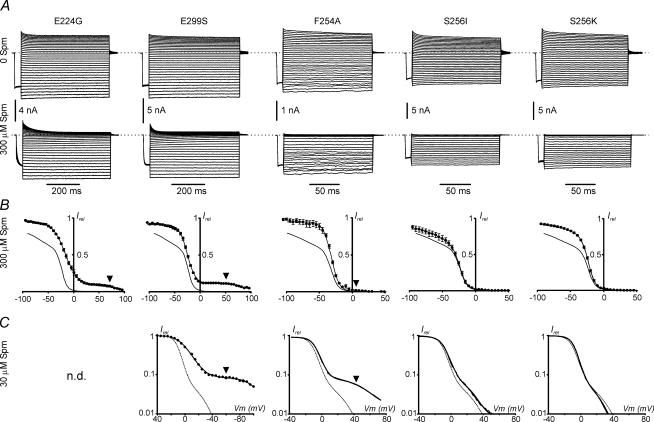
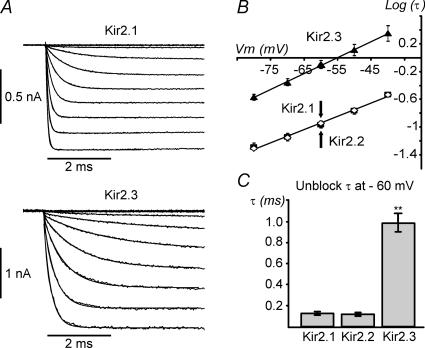
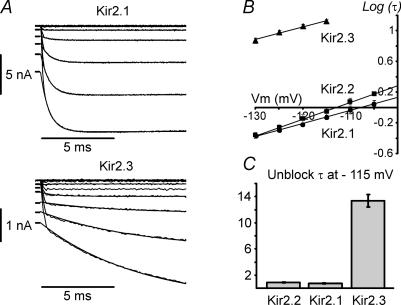
References
-
- Dhamoon AS, Jalife J. The inward rectifier current (IK1) controls cardiac excitability and is involved in arrhythmogenesis. Heart Rhythm. 2005;2:316–324. - PubMed
-
- Dhamoon AS, Pandit SV, Sarmast F, Parisian KR, Guha P, Li Y, Bagwe S, Taffet SM, Anumonwo JM. Unique Kir2.x properties determine regional and species differences in the cardiac inward rectifier K+ current. Circ Res. 2004;94:1332–1339. - PubMed
-
- Fakler B, Brandle U, Glowatzki E, Weidemann S, Zenner HP, Ruppersberg JP. Strong voltage-dependent inward rectification of inward rectifier K+ channels is caused by intracellular spermine. Cell. 1995;80:149–154. - PubMed
-
- Ficker E, Taglialatela M, Wible BA, Henley CM, Brown AM. Spermine and spermidine as gating molecules for inward rectifier K+ channels. Science. 1994;266:1068–1072. - PubMed