

Enthalpic and entropic contributions mediate the role of disulfide bonds on the conformational stability of interleukin-4
- PMID: 16373475
- PMCID: PMC2242368
- DOI: 10.1110/ps.051593306
Enthalpic and entropic contributions mediate the role of disulfide bonds on the conformational stability of interleukin-4
Abstract
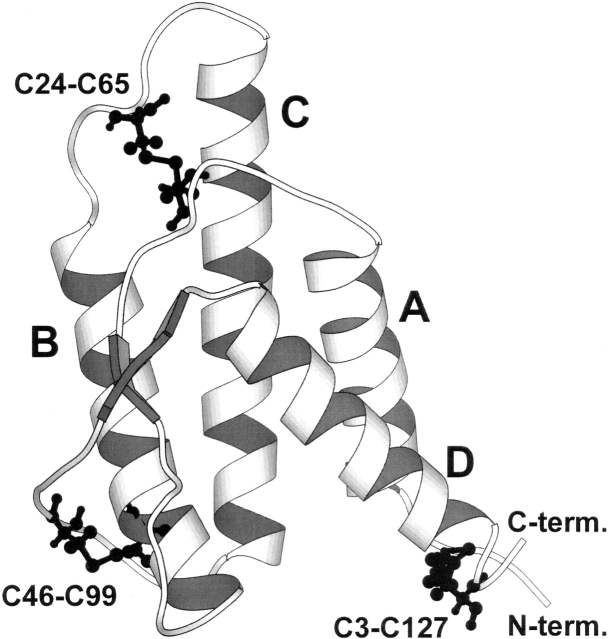
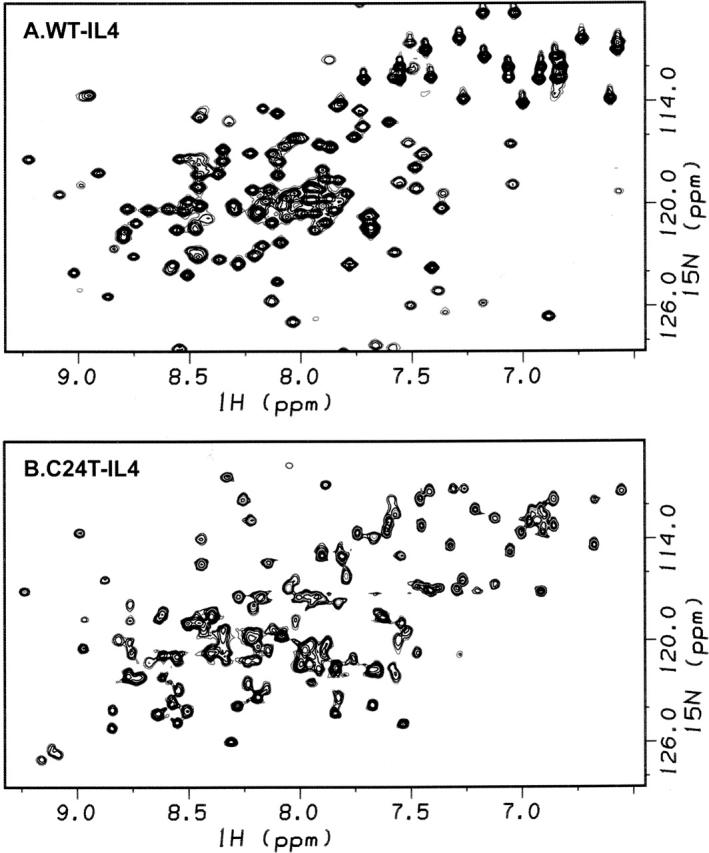
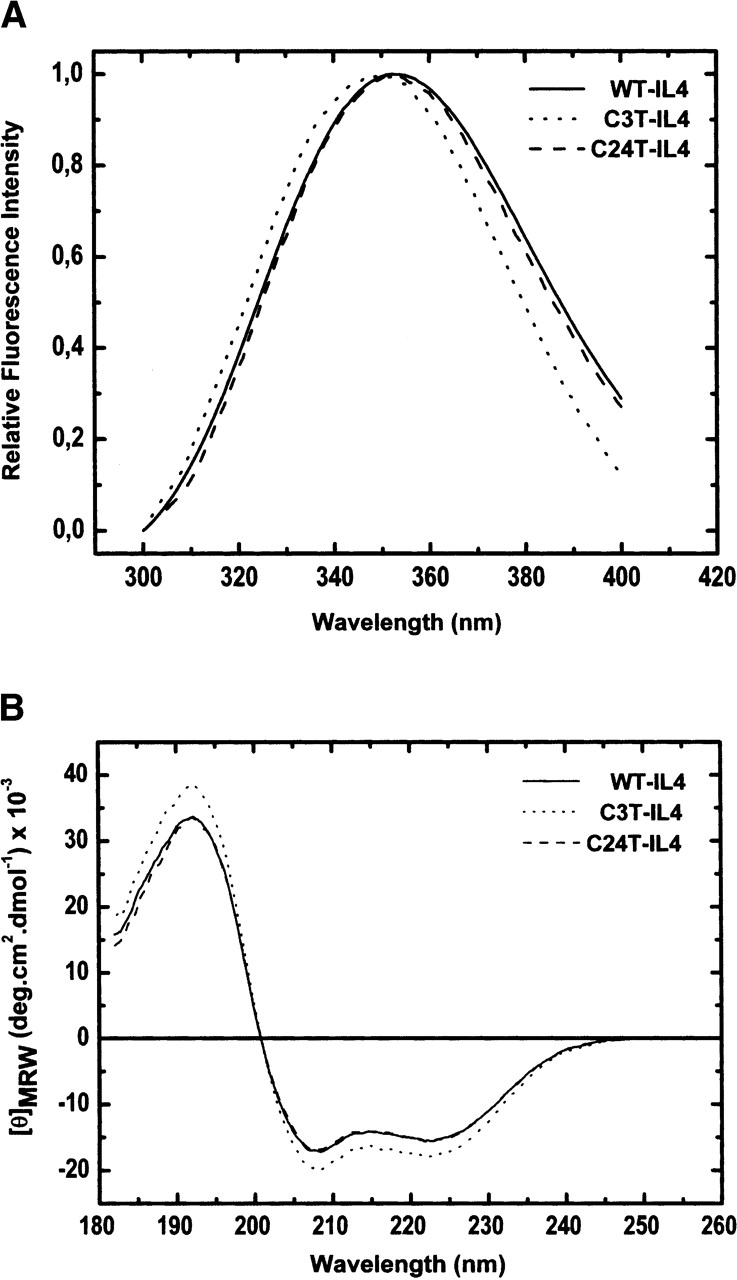
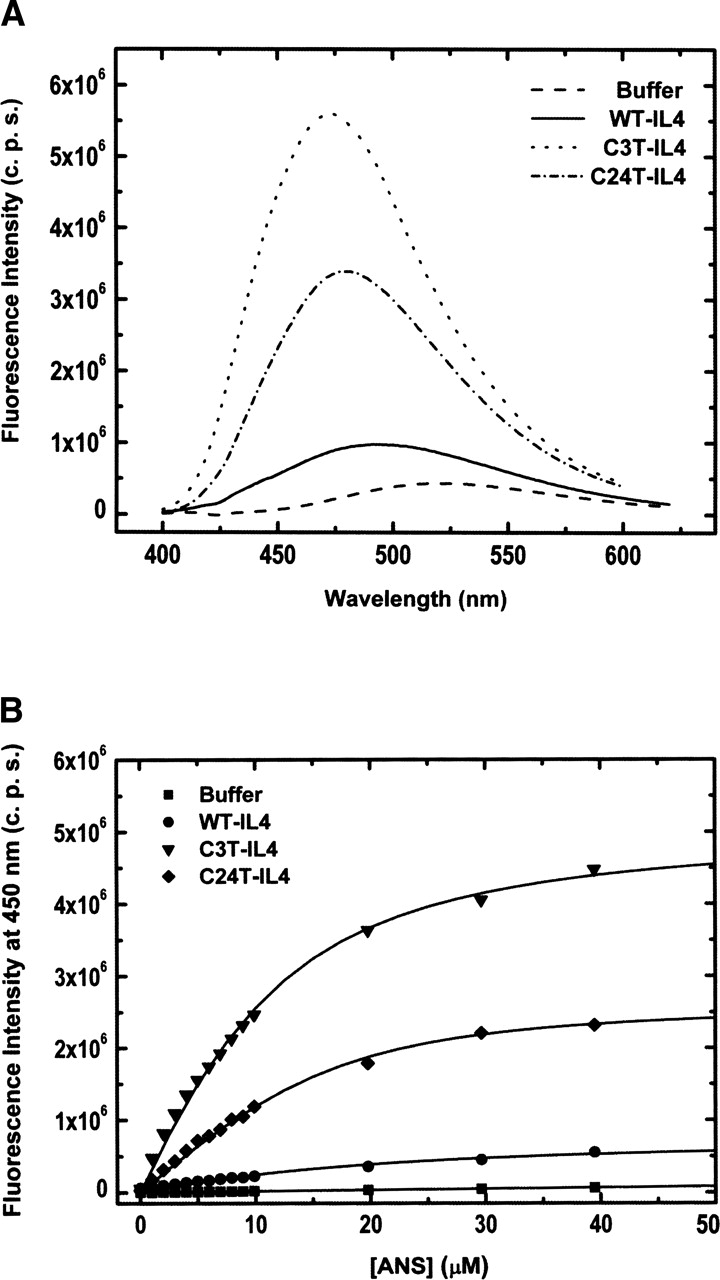
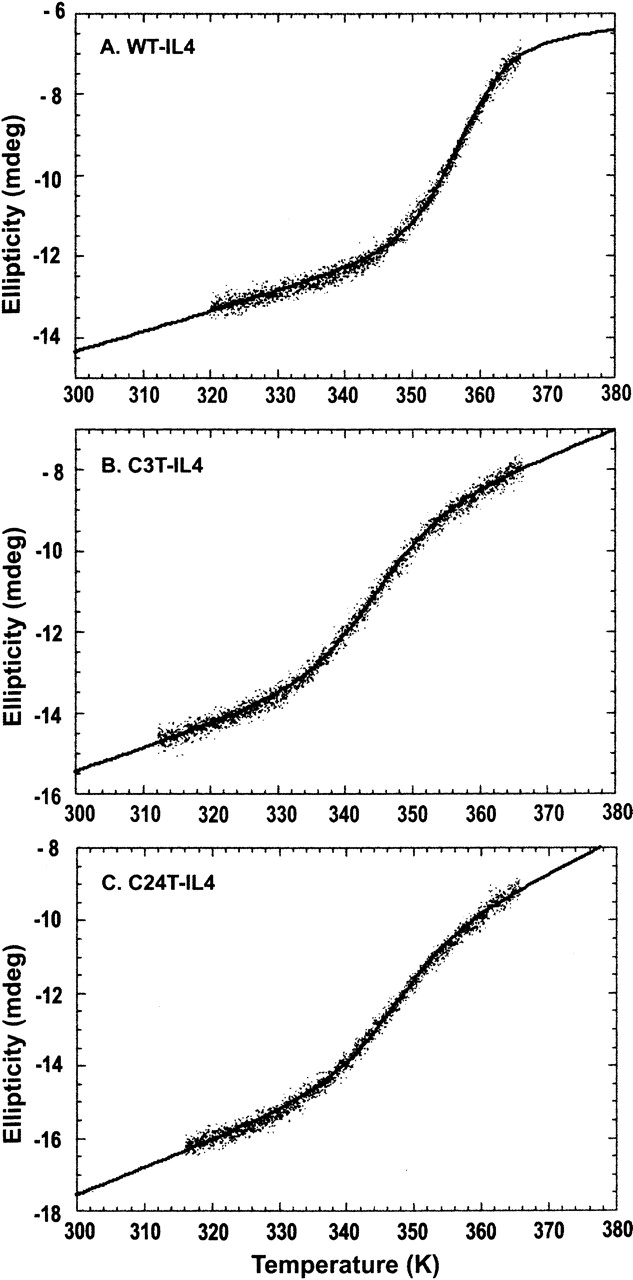
The role of disulfide bridges in the structure, stability, and folding pathways of proteins has been the subject of wide interest in the fields of protein design and engineering. However, the relative importance of entropic and enthalpic contributions for the stabilization of proteins provided by disulfides is not always clear. Here, we perform a detailed analysis of the role of disulfides in the conformational stability of human Interleukin-4 (IL4), a four-helix bundle protein. In order to evaluate the contribution of two out of the three disulfides to the structure and stability of IL4, two IL4 mutants, C3T-IL4 and C24T-IL4, were used. NMR and ANS binding experiments were compatible with altered dynamics and an increase of the nonpolar solvent-accessible surface area of the folded state of the mutant proteins. Chemical and thermal unfolding experiments followed by fluorescence and circular dichroism revealed that both mutant proteins have lower conformational stability than the wild-type protein. Transition temperatures of unfolding decreased 14 degrees C for C3T-IL4 and 10 degrees C for C24T-IL4, when compared to WT-IL4, and the conformational stability, at 25 degrees C, decreased 4.9 kcal/mol for C3T-IL4 and 3.2 kcal/mol for C24T-IL4. Interestingly, both the enthalpy and the entropy of unfolding, at the transition temperature, decreased in the mutant proteins. Moreover, a smaller change in heat capacity of unfolding was also observed for the mutants. Thus, disulfide bridges in IL4 play a critical role in maintaining the thermodynamic stability and core packing of the helix bundle.
Figures


References
-
- Alonso, D.O.V. and Dill, K.A. 1991. Solvent denaturation and stabilization of globular proteins. Biochemistry 30: 5974–5985. - PubMed
-
- Beldarrain, A., Lopez-Lacomba, J.L., Furrazola, G., Barberia, D., and Cortijo, M. 1999. Thermal denaturation of human γ-interferon. A calorimetric and spectroscopic study. Biochemistry 38: 7865–7873. - PubMed
-
- Bishop, B., Koay, D.C., Sartorelli, A.C., and Regan, L. 2001. Reengineering granulocyte colony-stimulating factor for enhanced stability. J. Biol. Chem. 276: 33465–33470. - PubMed
-
- Brems, D.N., Brown, P.L., and Becker, G.W. 1990. Equilibrium denaturation of human growth hormone and its cysteine-modified forms. J. Biol. Chem. 265: 5504–5511. - PubMed
-
- Cooper, A., Eyles, S.J., Radford, S.E., and Dobson, C.M. 1992. Thermo-dynamic consequences of the removal of a disulphide bridge from hen lysozyme. J. Mol. Biol. 225: 939–943. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
