

Purified hematopoietic stem cell engraftment of rare niches corrects severe lymphoid deficiencies without host conditioning
- PMID: 16380511
- PMCID: PMC2118067
- DOI: 10.1084/jem.20051714
Purified hematopoietic stem cell engraftment of rare niches corrects severe lymphoid deficiencies without host conditioning
Abstract
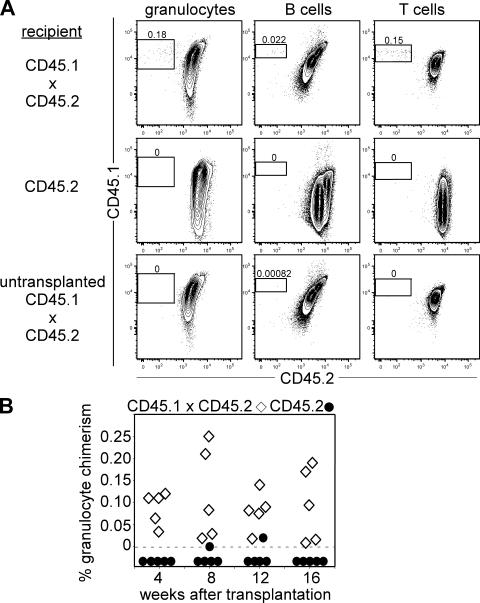
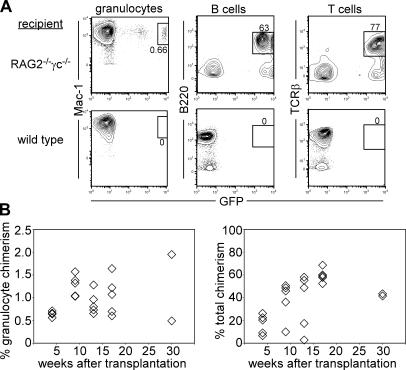
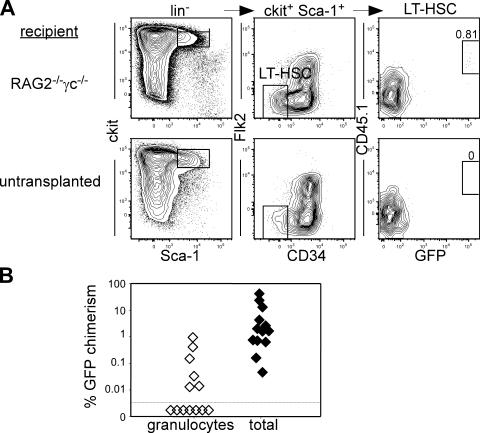
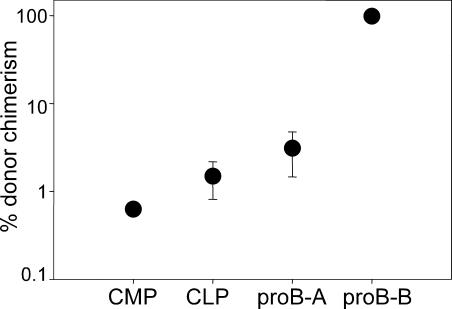
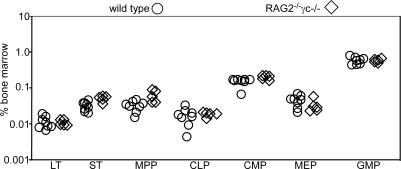
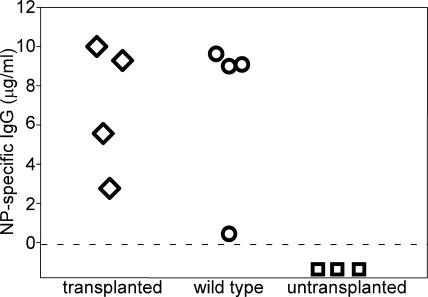
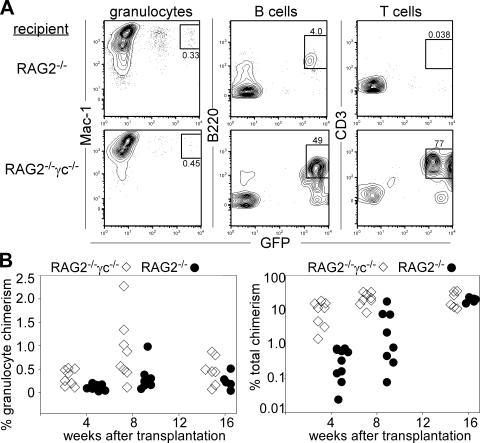
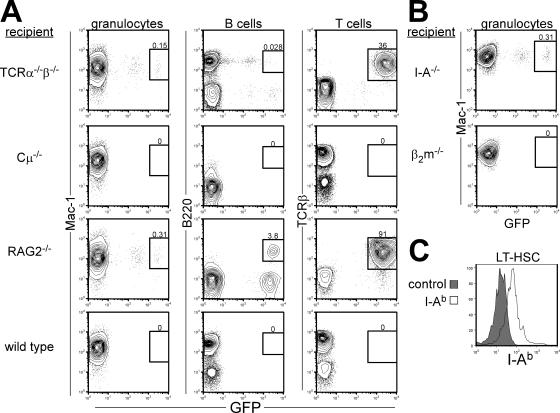
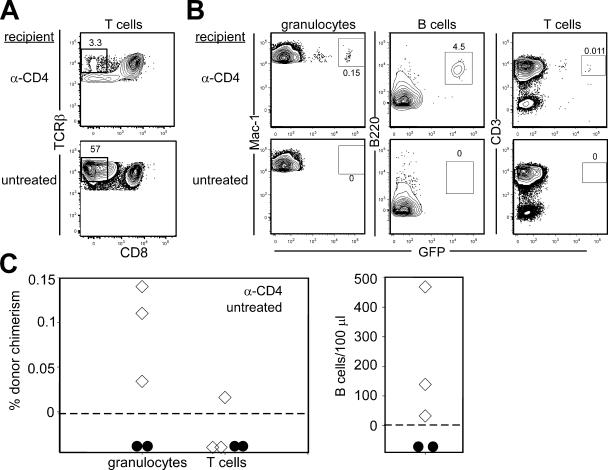
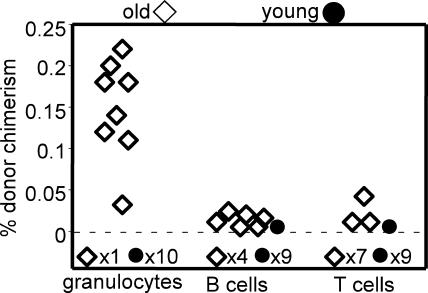
In the absence of irradiation or other cytoreductive conditioning, endogenous hematopoietic stem cells (HSCs) are thought to fill the unique niches within the bone marrow that allow maintenance of full hematopoietic potential and thus prevent productive engraftment of transplanted donor HSCs. By transplantation of purified exogenous HSCs into unconditioned congenic histocompatible strains of mice, we show that approximately 0.1-1.0% of these HSC niches are available for engraftment at any given point and find no evidence that endogenous HSCs can be displaced from the niches they occupy. We demonstrate that productive engraftment of HSCs within these empty niches is inhibited by host CD4+ T cells that recognize very subtle minor histocompatibility differences. Strikingly, transplantation of purified HSCs into a panel of severe combined immunodeficient (SCID) mice leads to a rapid and complete rescue of lymphoid deficiencies through engraftment of these very rare niches and expansion of donor lymphoid progenitors. We further demonstrate that transient antibody-mediated depletion of CD4+ T cells allows short-term HSC engraftment and regeneration of B cells in a mouse model of B(-) non-SCID. These experiments provide a general mechanism by which transplanted HSCs can correct hematopoietic deficiencies without any host conditioning or with only highly specific and transient lymphoablation.
Figures
References
-
- Morrison, S.J., A.M. Wandycz, K. Akashi, A. Globerson, and I.L. Weissman. 1996. The aging of hematopoietic stem cells. Nat. Med. 2:1011–1016. - PubMed
-
- Schofield, R. 1978. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells. 4:7–25. - PubMed
-
- Zhang, C.C., and H.F. Lodish. 2004. Insulin-like growth factor 2 expressed in a novel fetal liver cell population is a growth factor for hematopoietic stem cells. Blood. 103:2513–2521. - PubMed
-
- Calvi, L.M., G.B. Adams, K.W. Weibrecht, J.M. Weber, D.P. Olson, M.C. Knight, R.P. Martin, E. Schipani, P. Divieti, F.R. Bringhurst, et al. 2003. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature. 425:841–846. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
