

A lever-arm rotation drives motility of the minus-end-directed kinesin Ncd
- PMID: 16382238
- PMCID: PMC2851630
- DOI: 10.1038/nature04320
A lever-arm rotation drives motility of the minus-end-directed kinesin Ncd
Abstract
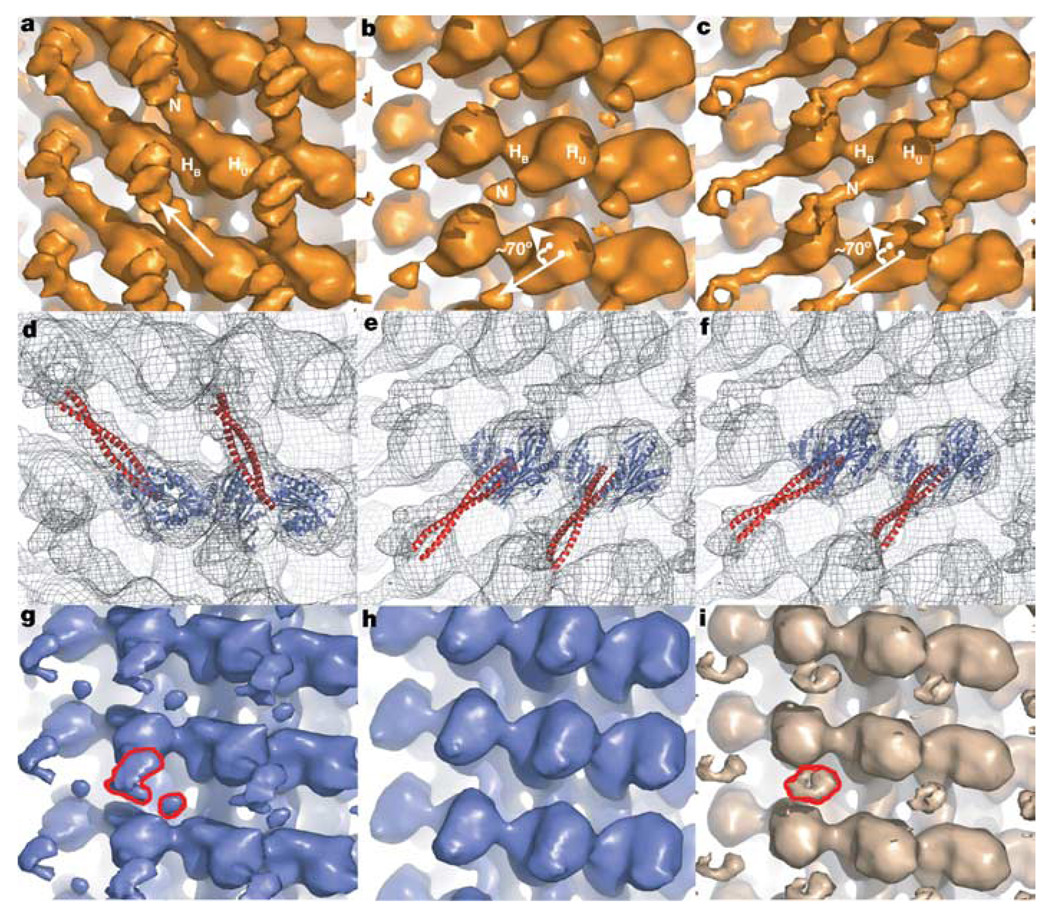
Kinesins are microtubule-based motor proteins that power intracellular transport. Most kinesin motors, exemplified by Kinesin-1, move towards the microtubule plus end, and the structural changes that govern this directional preference have been described. By contrast, the nature and timing of the structural changes underlying the minus-end-directed motility of Kinesin-14 motors (such as Drosophila Ncd) are less well understood. Using cryo-electron microscopy, here we demonstrate that a coiled-coil mechanical element of microtubule-bound Ncd rotates approximately 70 degrees towards the minus end upon ATP binding. Extending or shortening this coiled coil increases or decreases velocity, respectively, without affecting ATPase activity. An unusual Ncd mutant that lacks directional preference shows unstable nucleotide-dependent conformations of its coiled coil, underscoring the role of this mechanical element in motility. These results show that the force-producing conformational change in Ncd occurs on ATP binding, as in other kinesins, but involves the swing of a lever-arm mechanical element similar to that described for myosins.
Figures


Similar articles
-
A kinesin motor in a force-producing conformation.BMC Struct Biol. 2010 Jul 5;10:19. doi: 10.1186/1472-6807-10-19. BMC Struct Biol. 2010. PMID: 20602775 Free PMC article.
-
Kar3Vik1 uses a minus-end directed powerstroke for movement along microtubules.PLoS One. 2013;8(1):e53792. doi: 10.1371/journal.pone.0053792. Epub 2013 Jan 14. PLoS One. 2013. PMID: 23342004 Free PMC article.
-
Microscopic evidence for a minus-end-directed power stroke in the kinesin motor ncd.EMBO J. 2002 Nov 15;21(22):5969-78. doi: 10.1093/emboj/cdf622. EMBO J. 2002. PMID: 12426369 Free PMC article.
-
Kinesin motors as molecular machines.Bioessays. 2003 Dec;25(12):1212-9. doi: 10.1002/bies.10358. Bioessays. 2003. PMID: 14635256 Review.
-
New insights into the mechanochemical coupling mechanism of kinesin-microtubule complexes from their high-resolution structures.Biochem Soc Trans. 2023 Aug 31;51(4):1505-1520. doi: 10.1042/BST20221238. Biochem Soc Trans. 2023. PMID: 37560910 Free PMC article. Review.
Cited by
-
Anchoring geometry is a significant factor in determining the direction of kinesin-14 motility on microtubules.Sci Rep. 2022 Sep 14;12(1):15417. doi: 10.1038/s41598-022-19589-4. Sci Rep. 2022. PMID: 36104376 Free PMC article.
-
Motor Proteins.Cold Spring Harb Perspect Biol. 2018 May 1;10(5):a021931. doi: 10.1101/cshperspect.a021931. Cold Spring Harb Perspect Biol. 2018. PMID: 29716949 Free PMC article. Review.
-
Overview of the mechanism of cytoskeletal motors based on structure.Biophys Rev. 2018 Apr;10(2):571-581. doi: 10.1007/s12551-017-0368-1. Epub 2017 Dec 12. Biophys Rev. 2018. PMID: 29235081 Free PMC article. Review.
-
The ATPase pathway that drives the kinesin-14 Kar3Vik1 powerstroke.J Biol Chem. 2012 Oct 26;287(44):36673-82. doi: 10.1074/jbc.M112.395590. Epub 2012 Sep 12. J Biol Chem. 2012. PMID: 22977241 Free PMC article.
-
Bound ion effects: Using machine learning method to study the kinesin Ncd's binding with microtubule.Biophys J. 2024 Sep 3;123(17):2740-2748. doi: 10.1016/j.bpj.2023.12.024. Epub 2023 Dec 30. Biophys J. 2024. PMID: 38160255 Free PMC article.
References
-
- Hirokawa N, Takemura R. Kinesin superfamily proteins and their various functions and dynamics. Exp. Cell Res. 2004;301:50–59. - PubMed
-
- Sharp DJ, Rogers GC, Scholey JM. Microtubule motors in mitosis. Nature. 2000;407:41–47. - PubMed
-
- Rice S, et al. A structural change in the kinesin motor protein that drives motility. Nature. 1999;402:778–784. - PubMed
-
- Yildiz A, Tomishige M, Vale RD, Selvin PR. Kinesin walks hand-over-hand. Science. 2004;303:676–678. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
