

Review
doi: 10.1105/tpc.105.038513.
The structure of phytochrome: a picture is worth a thousand spectra
Affiliations
- PMID: 16387836
- PMCID: PMC1323480
- DOI: 10.1105/tpc.105.038513
Item in Clipboard
Review
The structure of phytochrome: a picture is worth a thousand spectra
Plant Cell.
2006 Jan.
Free PMC article
No abstract available
Figures

Bilin Chromophores, Photoconversion, and Domain Architecture of Phytochromes. (A) The bilin chromophores derived from phycocyanobilin (PCB), used by cyanobacterial and algal phytochromes (right), phytochromobilin (PΦB), used by plant phytochromes (right), and biliverdin (BV), used by the BphP subfamily (left), are shown along with their respective thioether linkages to the phytochrome apoproteins. Ring names (bold) and carbon numbering are indicated. (B) The photoconversion between Pr and Pfr phytochromes is shown along with the primary photochemical intermediates. The conversions of lumi-R into Pr and lumi-F into Pfr are thermal processes that lead to changes in the protein moieties that initiate signal output. (C) Conserved domains in the phytochrome primary sequence are shown (see text for description of PAS, GAF, and PHY domains). PAS and GAF domains are red and blue, respectively. The fragment of DrBphP whose structure has been reported (Wagner et al., 2005; PDB code 1ZTU) is shown at bottom with the positions of key chromophore-interacting residues shown in the primary sequence. HKRD, histidine kinase–related domain.
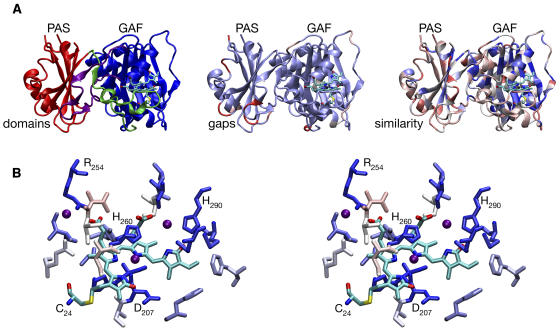
Conservation of Structural Elements among Phytochromes. (A) The DrBphP crystal structure (Wagner et al., 2005) is shown colored by domains (left), gaps (center), and similarity with other phytochromes (right). The BV chromophore and Cys24 (the site of covalent attachment in DrBphP) are shown colored by atom type (cyan, C; blue, N; red, O; yellow, S). Domains (left) are colored red (PAS domain), blue (GAF domain), green (N-terminal to the PAS domain, including part of the trefoil knot), and purple (lasso sequence inserted to the GAF domain and forming the remainder of the trefoil knot). Gaps (center) are colored with a continuous scale ranging from light blue (no gaps) to red (gaps ≥5 amino acids long) using an alignment of 122 members of the phytochrome family, including DrBphP. Similarity (right) is colored with a continuous scale ranging from dark blue (complete conservation) to red (variable) using a normalized BLOSUM62 substitution matrix (Henikoff and Henikoff, 1992) and the same sequence alignment. (B) Stereo view of chromophore binding in DrBphP. Cys24 and the BV chromophore are colored by element as in (A). Other residues within 4 Å of chromophore are colored by similarity as in (A). Water molecules within 4 Å of the BV chromophore are shown as purple spheres. Figures 2 to 4 were prepared with VMD, Tachyon, STRIDE, and homolmapper (Frishman and Argos, 1995; Humphrey et al., 1996; Stone, 1998).

The Trefoil Knot Interface between the PAS and GAF Domains. (A) A stereo view of the conserved knot core is shown colored as in Figure 2A (left). Amino acids 27 to 38 (green) are upstream of the PAS domain (red) and form the knot with amino acids 228 to 256 (the lasso, purple) that are inserted between β4 and α4 of the GAF domain (blue). (B) A simplified illustration of the trefoil knot topology is shown with colors as in (A). The GAF domain is formed by sequences on either side of the lasso, and residues that were not experimentally resolved in the loop between the PAS and GAF domains (Wagner et al., 2005) are shown as a dashed line. This schematic emphasizes the trefoil topology of the knot. (C) The interaction between the knot region and the chromophore is shown in stereo. The protein backbone is shown as a transparent trace colored as in (A), with residues 24 to 38 shown in green and Cys24, Arg254, and BV shown colored by atom type as in Figure 2.

Stereo Views of Interactions between the BV Chromophore and Nearby Residues in the DrBphP Apoprotein. (A) Residues within 4.5 Å of the chromophore D-ring are shown colored by atom type and as a single solvent-accessible surface prepared with MSMS (Sanner et al., 1996) colored by similarity as in Figure 2. Cys24 and the BV chromophore are shown in purple. The color highlighting of the protein residues ranges from dark blue (absolutely conserved) to light blue (strongly conserved). No red-shaded (variable) residues are located near the chromophore. (B) Tight packing of His260, Ile208, and Tyr216 about C10 is shown with Cys24 and chromophore in purple. Amino acid residues are shown colored by atom type as in Figure 2A.

Structures of Synthetically Locked Bilins and Spectra of Their Holophytochrome Adducts. Wild-type BV (top panels) or two synthetic analogs locked in either the C15-Z,anti (middle panels) or C15-E,anti (bottom panels) configuration are shown assembled with the bacteriophytochrome Agp1. Absorption spectra of the resulting adducts are shown on the left, and structures of the bilin chromophores are shown on the right. Atoms determining the configuration about C15 are highlighted in blue. The wild-type BV adduct is shown in the Pr state (solid) and as a calculated pure Pfr spectrum (dashed line). Figures were prepared from raw data kindly provided by Tilman Lamparter and Katsuhiko Inomata (Inomata et al., 2005).
References
-
- Andel, F., Lagarias, J.C., and Mathies, R.A. (1996). Resonance Raman analysis of chromophore structure in the lumi-R photoproduct of phytochrome. Biochemistry 35 15997–16008. - PubMed
-
- Andel, F., Murphy, J.T., Haas, J.A., McDowell, M.T., van der Hoef, I., Lugtenburg, J., Lagarias, J.C., and Mathies, R.A. (2000). Probing the photoreaction mechanism of phytochrome through analysis of resonance Raman vibrational spectra of recombinant analogues. Biochemistry 39 2667–2676. - PubMed
-
- Aravind, L., and Ponting, C.P. (1997). The GAF domain: An evolutionary link between diverse phototransducing proteins. Trends Biochem. Sci. 22 458–459. - PubMed
-
- Batschauer, A. (2003). Photoreceptors and Light Signaling. (Cambridge, UK: Royal Society of Chemistry).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
