

PKA-mediated phosphorylation regulates the function of activation-induced deaminase (AID) in B cells
- PMID: 16387847
- PMCID: PMC1326186
- DOI: 10.1073/pnas.0509969103
PKA-mediated phosphorylation regulates the function of activation-induced deaminase (AID) in B cells
Abstract
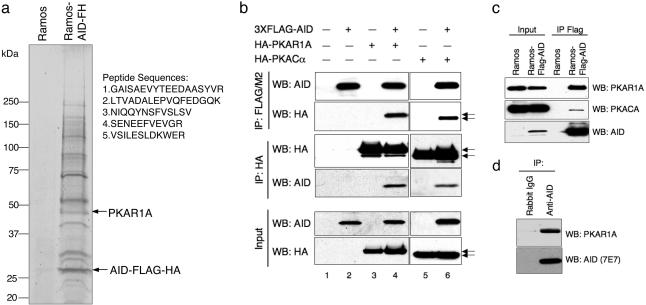
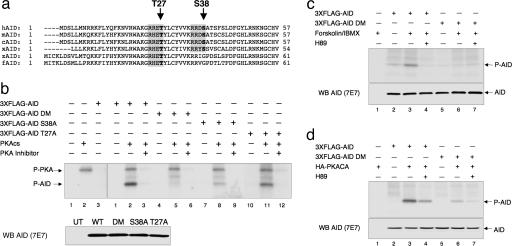
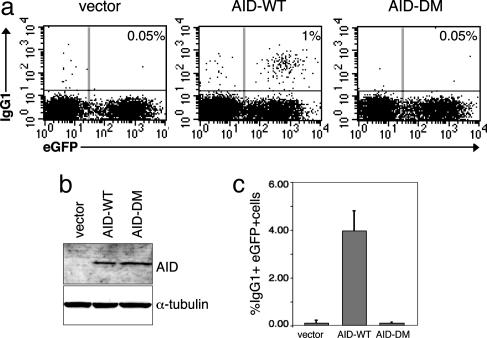
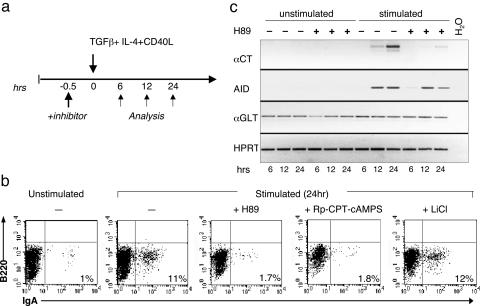
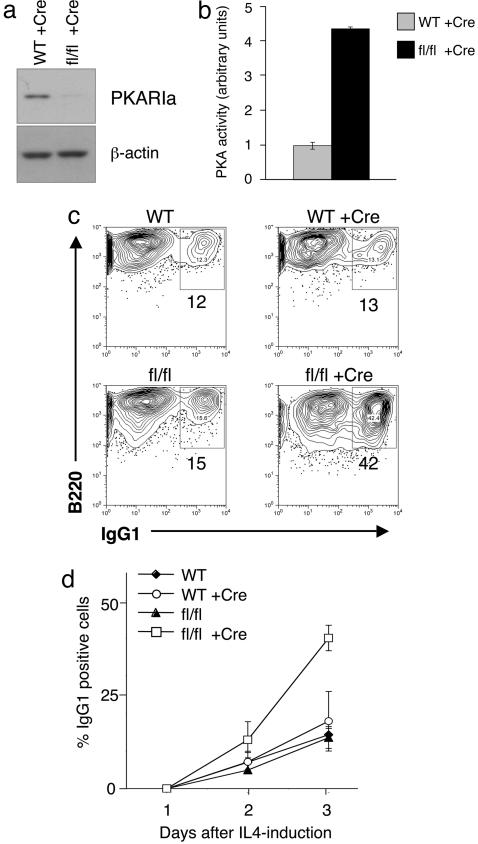
During humoral immune responses, two distinct genetic modification events diversify the Ig genes in germinal center (GC) B cells: somatic hypermutation and class switch recombination (CSR). Both processes require the activity of activation-induced cytidine deaminase (AID), an enzyme expressed specifically in GC B cells. However, the mechanisms that regulate AID activity are largely unknown. Here we report that protein kinase A (PKA) phosphorylates AID and regulates its activity in GC B cells. AID physically interacts with the PKA holoenzyme in the cytoplasm and is phosphorylated by the PKA catalytic subunit at specific residues. AID phosphorylation is required for CSR, because substitution of the two phosphorylation targets impairs its ability to rescue CSR in AID-deficient B cells. Pharmacologic inhibition of PKA prevents isotype class switching in a murine B-cell lymphoma cell line; conversely, B cells from mice where PKA activity is made constitutive by conditional deletion of the PKA regulatory subunit gene display enhanced CSR. These findings implicate PKA in the regulation of AID function and suggest that the control of T cell-dependent immune responses may be modulated, via AID, by signals that activate PKA.
Figures
References
-
- Rajewsky, K. (1996) Nature 381, 751-758. - PubMed
-
- Papavasiliou, F. N. & Schatz, D. G. (2002) Cell 109, Suppl., S35-S44. - PubMed
-
- Manis, J. P., Tian, M. & Alt, F. W. (2002) Trends Immunol. 23, 31-39. - PubMed
-
- Honjo, T., Kinoshita, K. & Muramatsu, M. (2002) Annu. Rev. Immunol. 20, 165-196. - PubMed
-
- Revy, P., Muto, T., Levy, Y., Geissmann, F., Plebani, A., Sanal, O., Catalan, N., Forveille, M., Dufourcq-Labelouse, R., Gennery, A., et al. (2000) Cell 102, 565-575. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
