

A new approach to producing functional G alpha subunits yields the activated and deactivated structures of G alpha(12/13) proteins
- PMID: 16388592
- PMCID: PMC2688741
- DOI: 10.1021/bi051729t
A new approach to producing functional G alpha subunits yields the activated and deactivated structures of G alpha(12/13) proteins
Abstract
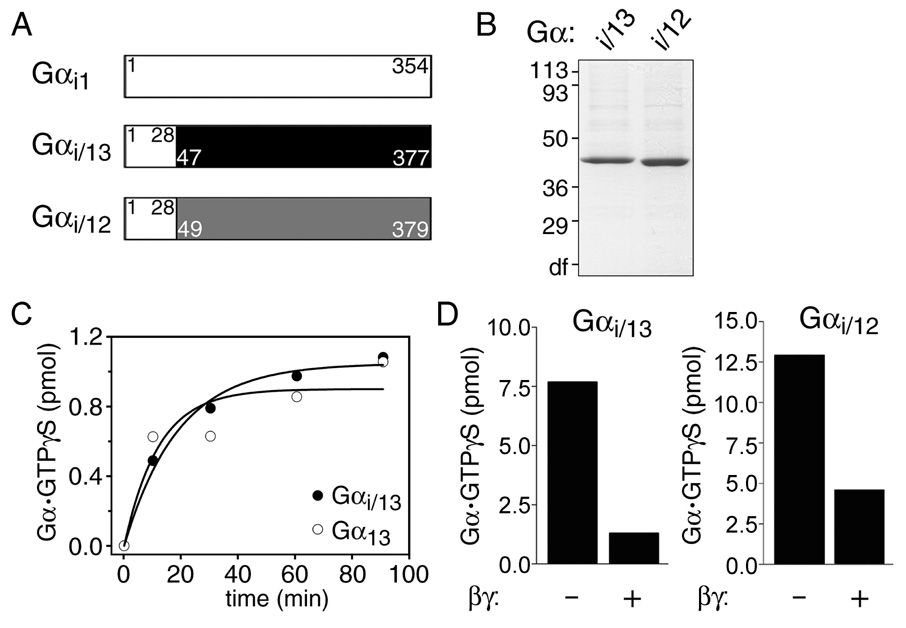
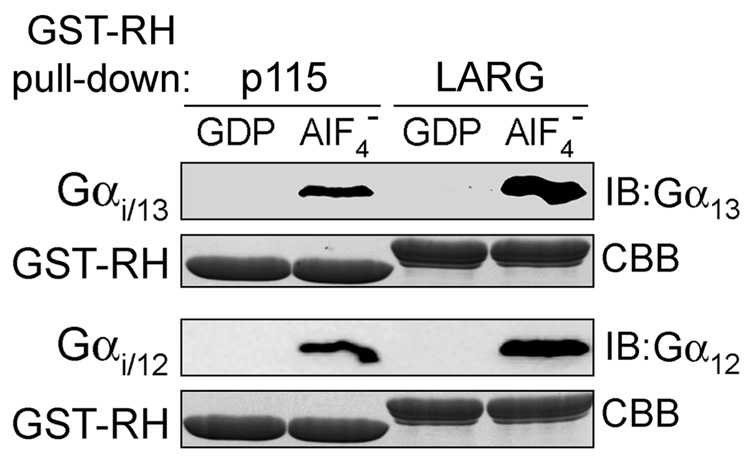
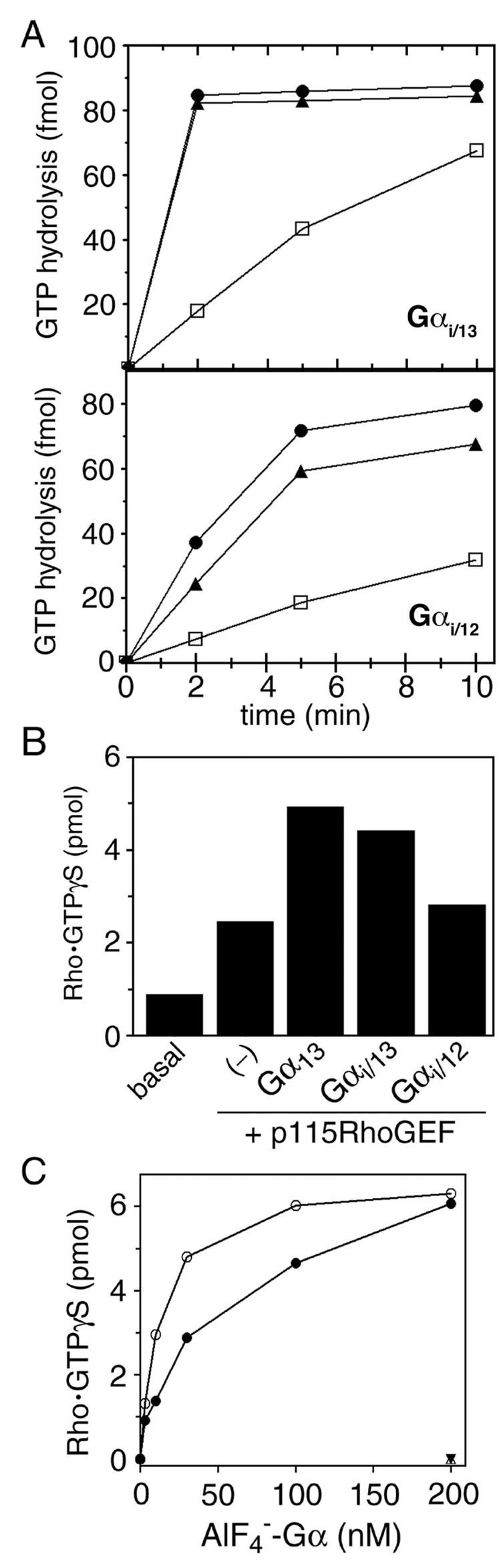
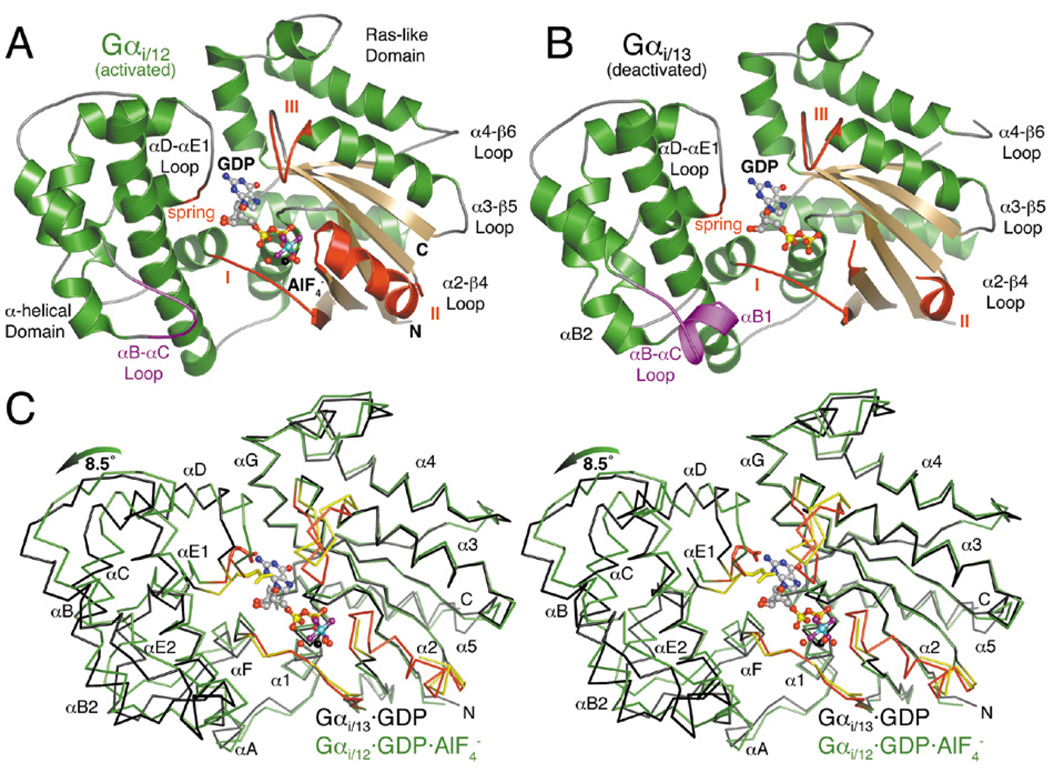
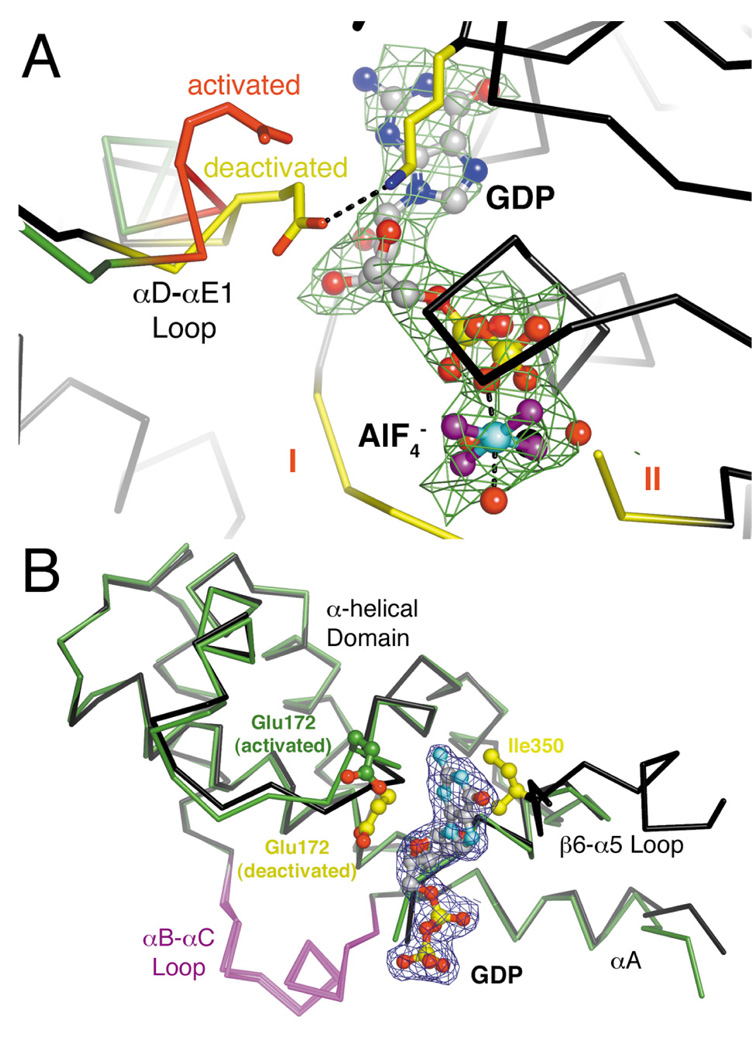
The oncogenic G(12/13) subfamily of heterotrimeric G proteins transduces extracellular signals that regulate the actin cytoskeleton, cell cycle progression, and gene transcription. Previously, structural analyses of fully functional G alpha(12/13) subunits have been hindered by insufficient amounts of homogeneous, functional protein. Herein, we report that substitution of the N-terminal helix of G alpha(i1) for the corresponding region of G alpha12 or G alpha13 generated soluble chimeric subunits (G alpha(i/12) and G alpha(i/13)) that could be purified in sufficient amounts for crystallographic studies. Each chimera bound guanine nucleotides, G betagamma subunits, and effector proteins and exhibited GAP responses to p115RhoGEF and leukemia-associated RhoGEF. Like their wild-type counterparts, G alpha(i/13), but not G alpha(i/12), stimulated the activity of p115RhoGEF. Crystal structures of the G alpha(i/12) x GDP x AlF4(-) and G alpha(i/13) x GDP complexes were determined using diffraction data extending to 2.9 and 2.0 A, respectively. These structures reveal not only the native structural features of G alpha12 and G alpha13 subunits, which are expected to be important for their interactions with GPCRs and effectors such as G alpha-regulated RhoGEFs, but also novel conformational changes that are likely coupled to GTP hydrolysis in the G alpha(12/13) class of heterotrimeric G proteins.
Figures
References
-
- Neves SR, Ram PT, Iyengar R. G protein pathways. Science. 2002;296:1636–1639. - PubMed
-
- Hollinger S, Hepler JR. Cellular regulation of RGS proteins: modulators and integrators of G protein signaling. Pharmacol Rev. 2002;54:527–559. - PubMed
-
- Berman DM, Kozasa T, Gilman AG. The GTPase-activating protein RGS4 stabilizes the transition state for nucleotide hydrolysis. J Biol Chem. 1996;271:27209–27212. - PubMed
-
- Tesmer JJG, Berman DM, Gilman AG, Sprang SR. Structure of RGS4 bound to AlF4−-activated Giα1: stabilization of the transition state for GTP hydrolysis. Cell. 1997;89:251–261. - PubMed
-
- Hepler JR, Gilman AG. G proteins. Trends Biochem Sci. 1992;17:383–387. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
