

A mixture theory analysis for passive transport in osmotic loading of cells
- PMID: 16389086
- PMCID: PMC2859701
- DOI: 10.1016/j.jbiomech.2004.12.013
A mixture theory analysis for passive transport in osmotic loading of cells
Abstract
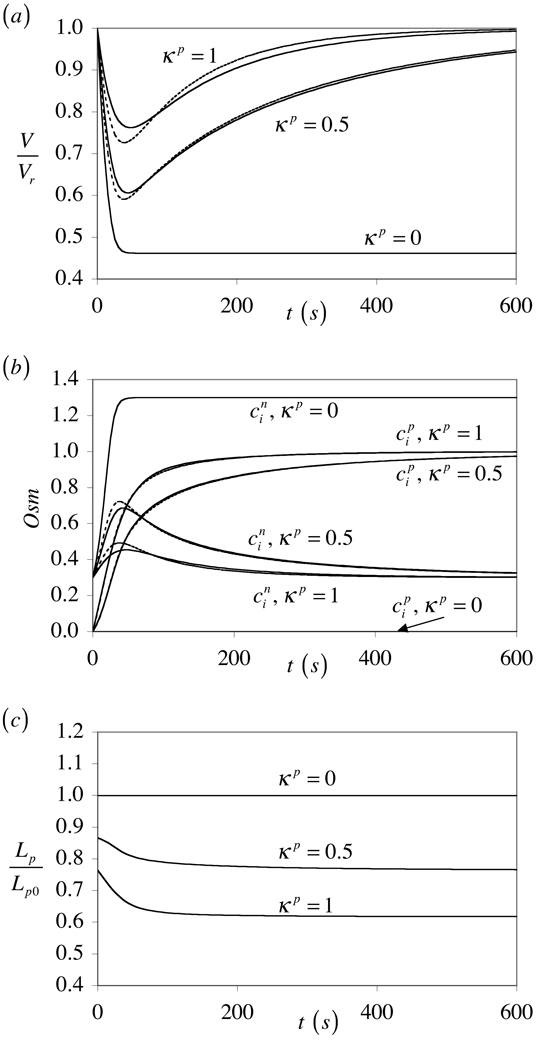
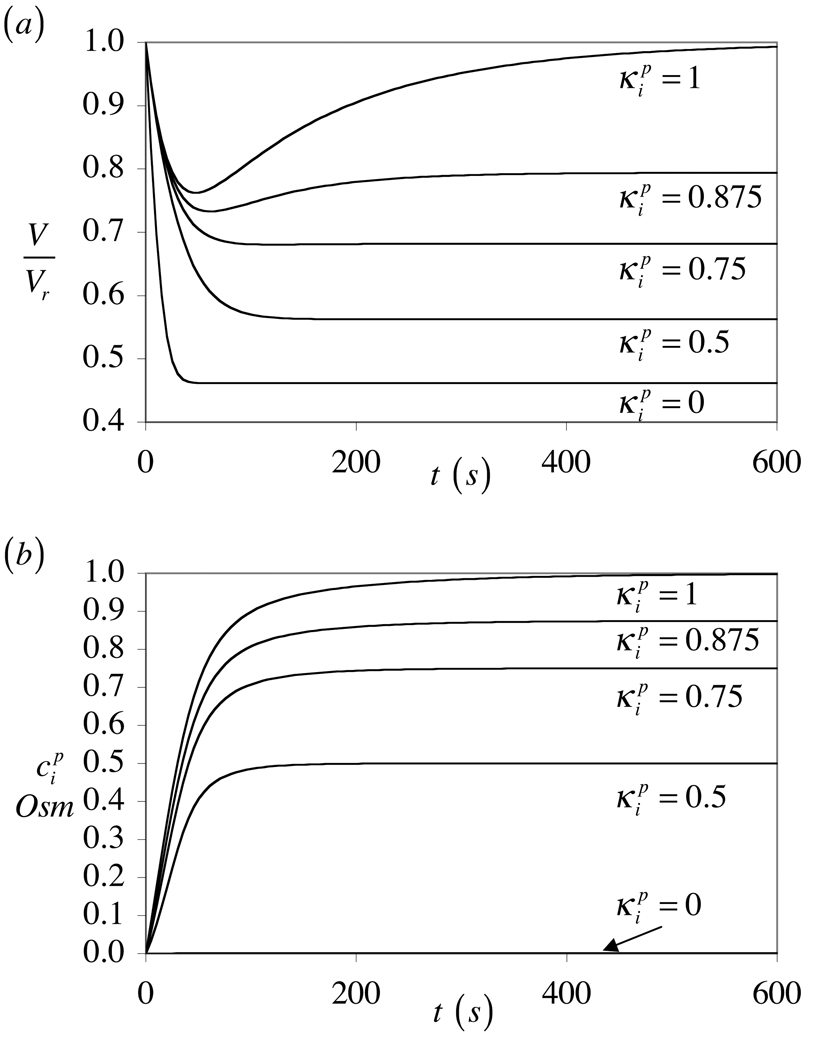
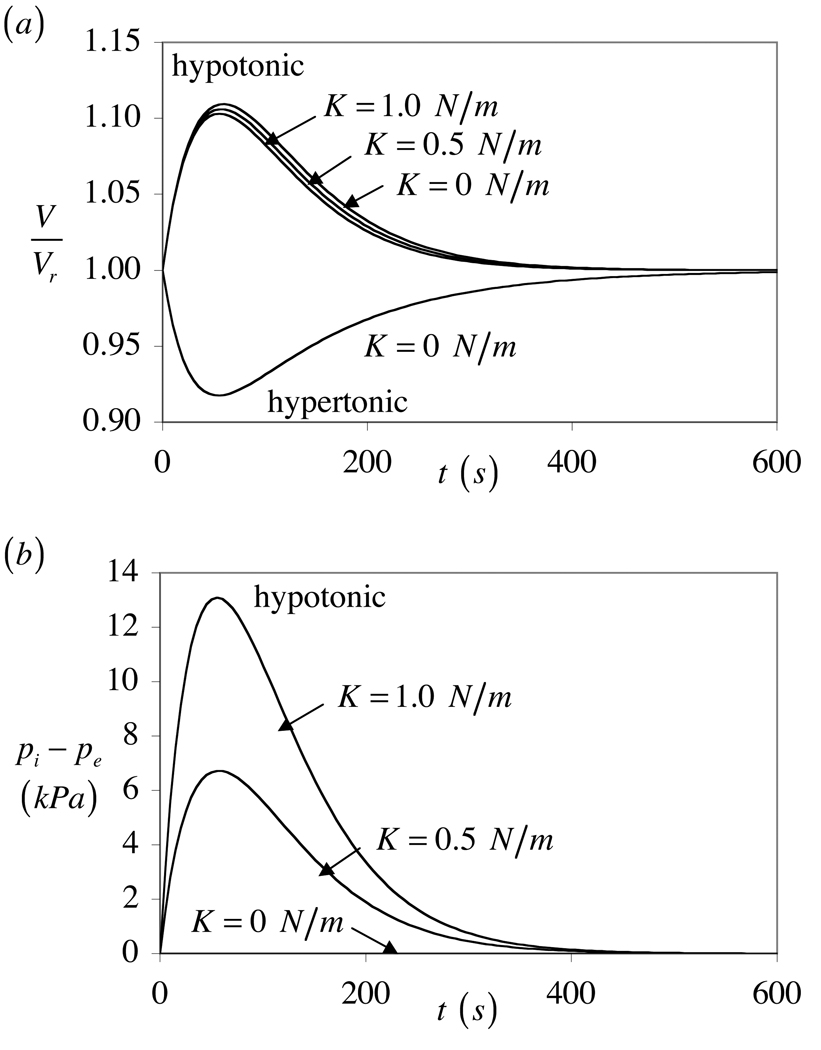
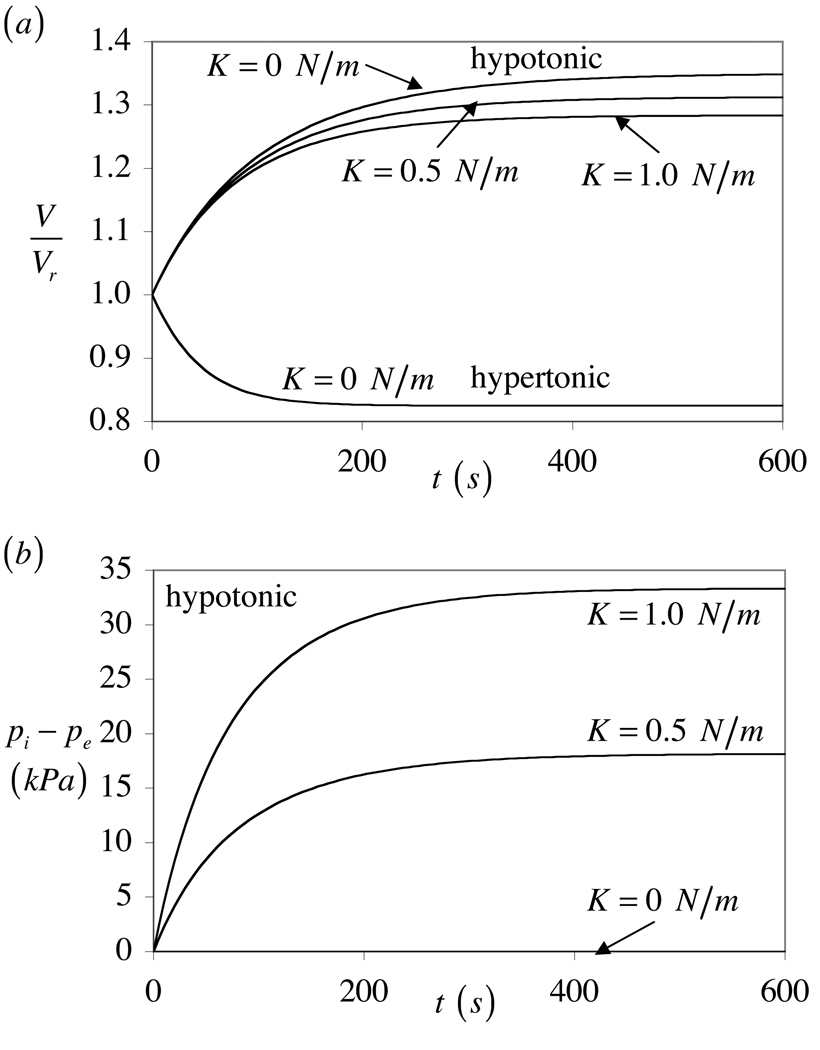
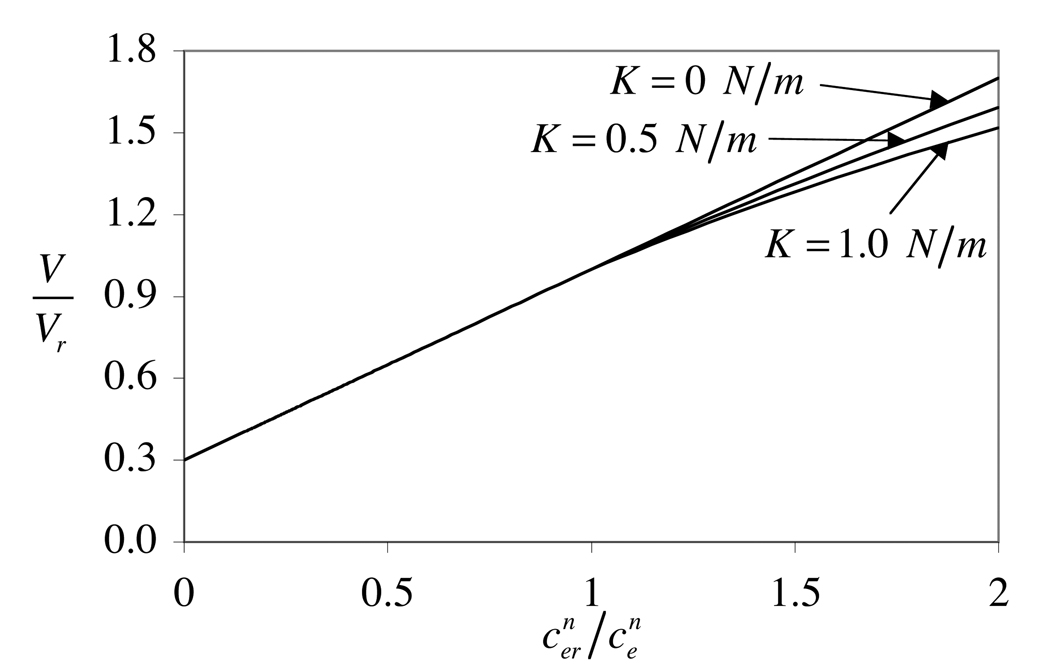
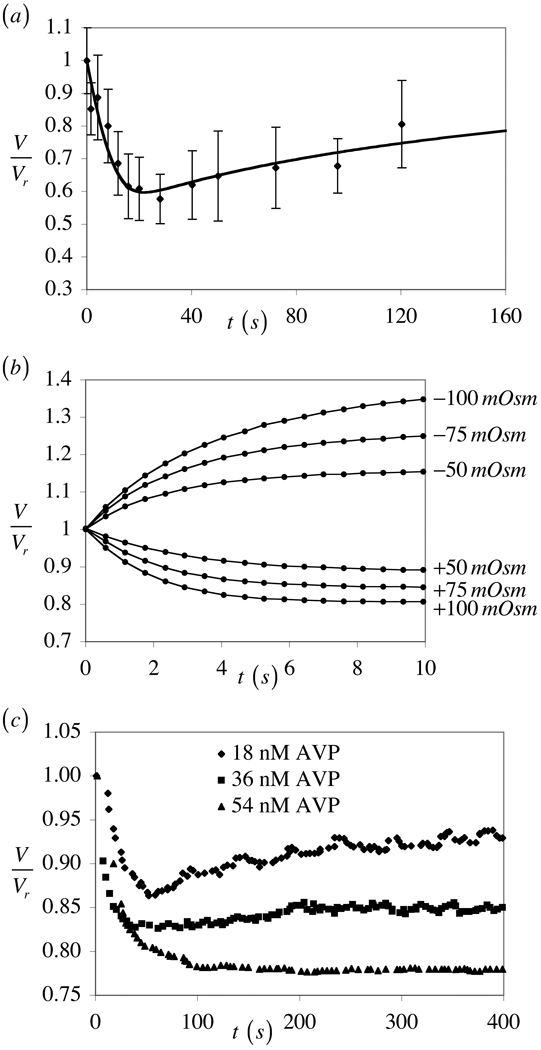
The theory of mixtures is applied to the analysis of the passive response of cells to osmotic loading with neutrally charged solutes. The formulation, which is derived for multiple solute species, incorporates partition coefficients for the solutes in the cytoplasm relative to the external solution, and accounts for cell membrane tension. The mixture formulation provides an explicit dependence of the hydraulic conductivity of the cell membrane on the concentration of permeating solutes. The resulting equations are shown to reduce to the classical equations of Kedem and Katchalsky in the limit when the membrane tension is equal to zero and the solute partition coefficient in the cytoplasm is equal to unity. Numerical simulations demonstrate that the concentration-dependence of the hydraulic conductivity is not negligible; the volume response to osmotic loading is very sensitive to the partition coefficient of the solute in the cytoplasm, which controls the magnitude of cell volume recovery; and the volume response is sensitive to the magnitude of cell membrane tension. Deviations of the Boyle-van't Hoff response from a straight line under hypo-osmotic loading may be indicative of cell membrane tension.
Figures
References
-
- Atkin RJ, Craine RE. Continuum Theories of Mixtures: Basic Theory and Historical Development. Q J Mech Appl Math. 1976;29:209–244.
-
- Bowen RM. Incompressible porous media models by use of the theory of mixtures. International Journal of Engineering Science. 1980;18:1129–1148.
-
- Conte SD, De Boor C. Elementary numerical analysis: an algorithmic approach. New York: McGraw-Hill; 1980.
-
- Curry MR, Kleinhans FW, Watson PF. Measurement of the water permeability of the membranes of boar, ram, and rabbit spermatozoa using concentration-dependent self-quenching of an entrapped fluorophore. Cryobiology. 2000;41:167–173. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
