

Lateral root initiation in Arabidopsis: developmental window, spatial patterning, density and predictability
- PMID: 16390845
- PMCID: PMC2803408
- DOI: 10.1093/aob/mcj604
Lateral root initiation in Arabidopsis: developmental window, spatial patterning, density and predictability
Erratum in
- Ann Bot (Lond). 2006 Nov;98(5):1115
Abstract
Background and aims: The basic regulatory mechanisms that control lateral root (LR) initiation are still poorly understood. An attempt is made to characterize the pattern and timing of LR initiation, to define a developmental window in which LR initiation takes place and to address the question of whether LR initiation is predictable.
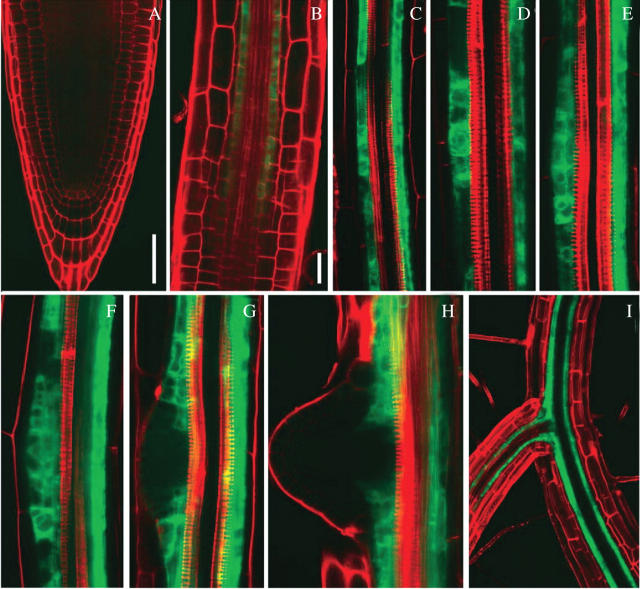
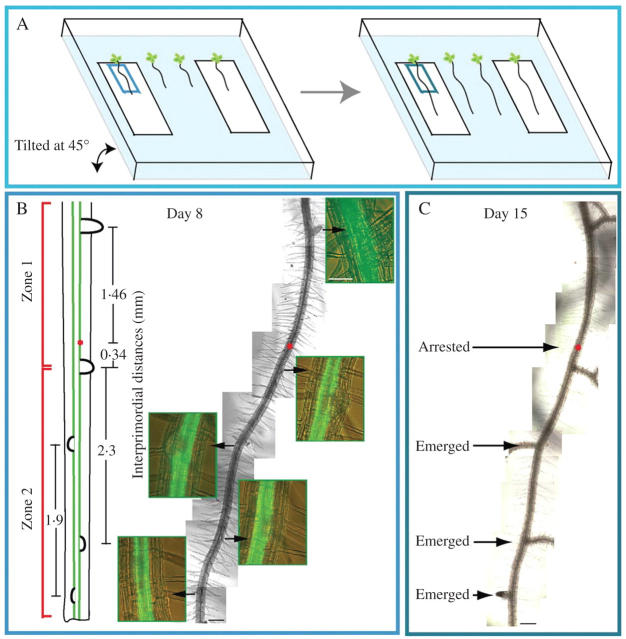
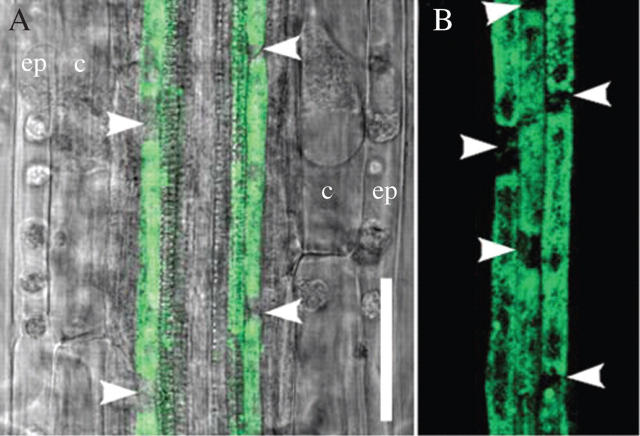
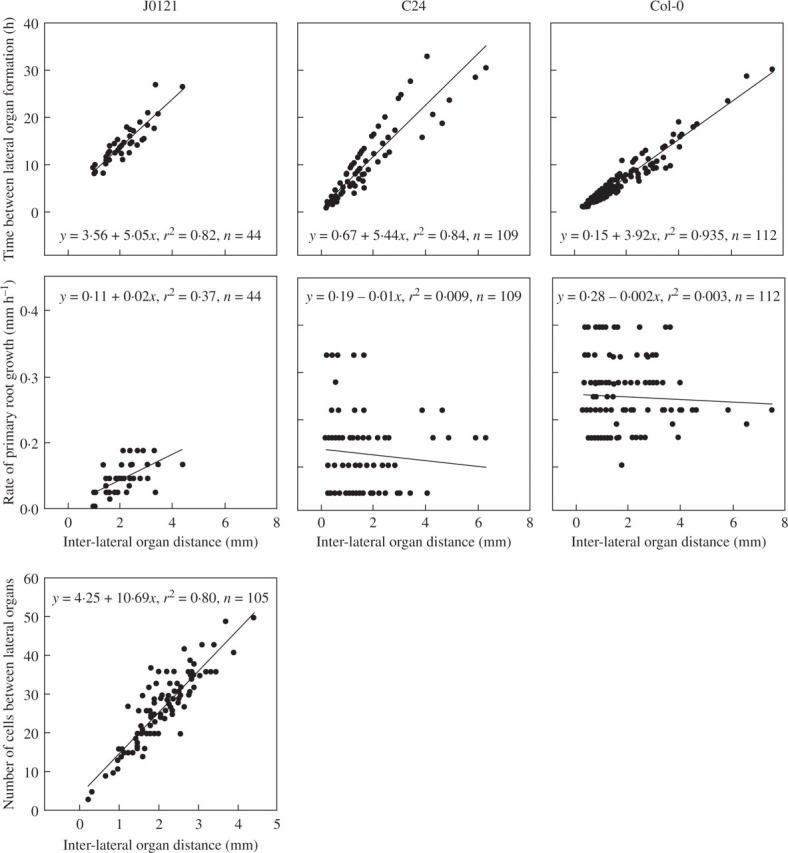
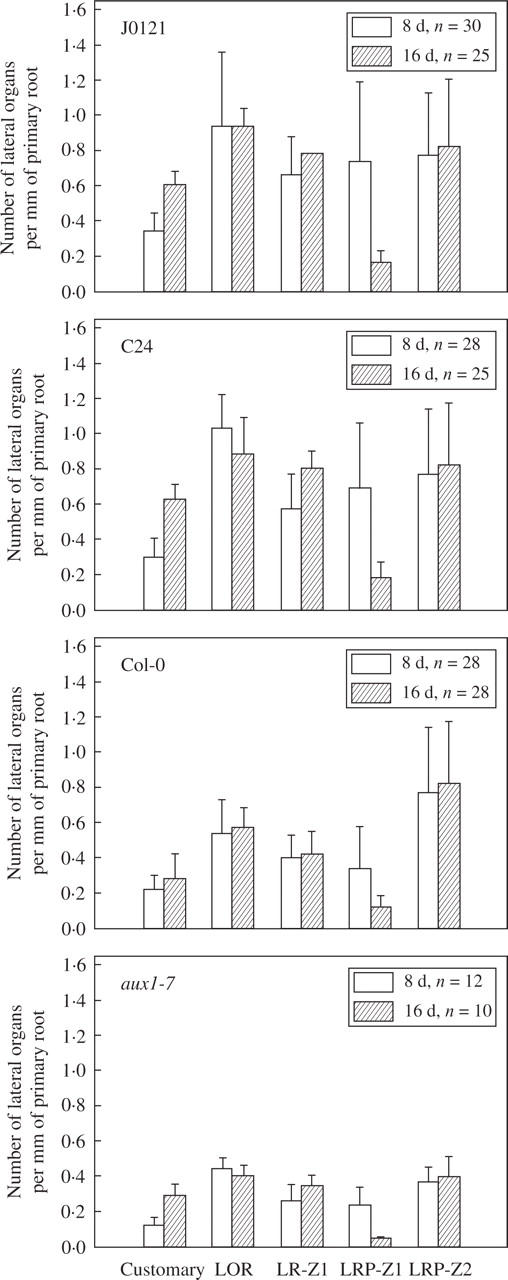
Methods: The spatial patterning of LRs and LR primordia (LRPs) on cleared root preparations were characterized. New measures of LR and LRP densities (number of LRs and/or LRPs divided by the length of the root portions where they are present) were introduced and illustrate the shortcomings of the more customarily used measure through a comparative analysis of the mutant aux1-7. The enhancer trap line J0121 was used to monitor LR initiation in time-lapse experiments and a plasmolysis-based method was developed to determine the number of pericycle cells between successive LRPs.
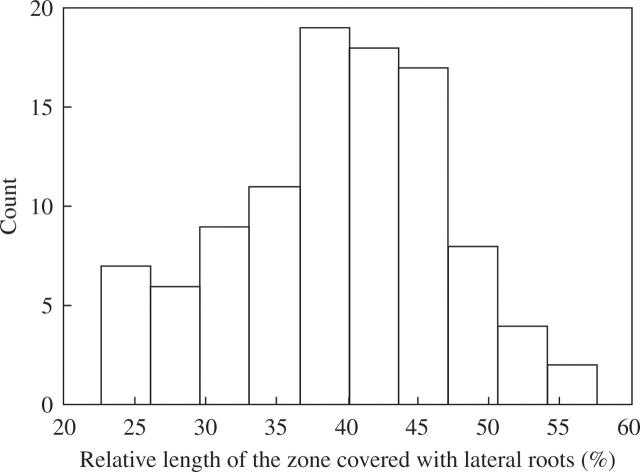
Key results: LRP initiation occurred strictly acropetally and no de novo initiation events were found between already developed LRs or LRPs. However, LRPs did not become LRs in a similar pattern. The longitudinal spacing of lateral organs was variable and the distance between lateral organs was proportional to the number of cells and the time between initiations of successive LRPs. There was a strong tendency towards alternation in LR initiation between the two pericycle cell files adjacent to the protoxylem poles. LR density increased with time due to the emergence of slowly developing LRPs and appears to be unique for individual Arabidopsis accessions.
Conclusions: In Arabidopsis there is a narrow developmental window for LR initiation, and no specific cell-count or distance-measuring mechanisms have been found that determine the site of successive initiation events. Nevertheless, the branching density and lateral organ density (density of LRs and LRPs) are accession-specific, and based on the latter density the average distance between successive LRs can be predicted.
Figures
References
-
- Aloni R, Langhans M, Aloni E, Dreieicher E, Ullrich CI. 2005. Root-synthesized cytokinin in Arabidopsis is distributed in the shoot by the transpiration stream. Journal of Experimental Botany 56: 1535–1544. - PubMed
-
- Barlow PW, Adam JS. 1988. The position and growth of lateral roots on cultured root axes of tomato, Lycopersicon esculentum (Solanaceae). Plant Systematics and Evolution 158: 141–154.
-
- Barlow PW, Volkmann D, Baluška F. 2004. Polarity in roots. In: Lindsey K, ed. Polarity in plants. Oxford: Blackwell Publishing, 192–241.
-
- Blakely LM, Durham M, Evans TA, Blakely RM. 1982. Experimental studies on lateral root formation in radish seedling roots. I. General methods, developmental stages, and spontaneous formation of laterals. Botanical Gazette 143: 341–352.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
